|
||||
|
|
ГЛАВА 3. ОТ ЭРЕКТУСОВ К САПИЕНСАМЭректусы осваивают мирВ течение второго миллионолетия до н. э. эректусы постепенно расселялись по просторам Евразии, неся с собой древнейшую технологию обработки камня — олдувайскую. Технические усовершенствования если и появлялись кое-где за пределами Африки, то редко и незначительные. Археологи полагают, что евразийские популяции эректусов были немногочисленны и сильно разобщены, хотя эпизодические встречи и обмен генами между ними, возможно, все-таки происходили, так что в масштабе сотен тысячелетий могла поддерживаться относительная генетическая общность человечества. Но разобщенность была достаточной для того, чтобы эректусы смогли подразделиться на несколько географических рас (что они, возможно, и сделали). В Африке плотность населения и разнообразие гоминид были куда выше, чем в Евразии. Здесь еще долго бок о бок с эректусами жили хабилисы и парантропы (и вовсе не факт, что представители некоторых популяций, относимых к разным видам, не могли иногда друг с другом скрещиваться) (К тому времени (1,9-1,5 млн лет назад) с момента разделения эволюционных линий хабилисов, эректусов и парантропов прошло, по-видимому, не более 1-2 млн лет. Этого недостаточно, чтобы считать полностью исключенной возможность межвидовой гибридизации. Обычно для достижения полной генетической несовместимости между разобщенными популяциями требуется в среднем около 3 млн лет. К тому же у приматов известны как межвидовые, так и межродовые гибриды: например, между геладами (Theropithecus gelada) и гамадрилами (Papio bamadryas). Судя по палеонтологическим данным, эти два рода разошлись более 3,5 млн лет назад (Jolly et al., 199/). Межродовые гибриды обычно бесплодны, но межвидовые (например, между разными видами макак) могут быть плодовитыми), да и самих эректусов было больше. В такой разнообразной и насыщенной среде эволюция шла быстрее. Вероятно, сказывались и более острая конкуренция — стимул для эволюционной "гонки вооружений", и возможности для культурного обмена. Проявилось это прежде всего в развитии каменных технологий. Олдувайская технология основана на производстве острых отщепов случайной формы, которые используются в качестве орудий, тогда как остающиеся "ядра" (нуклеусы), как правило, представляют собой производственные отходы. Около 1,7 млн лет назад в Африке появляются первые орудия нового, ашельского типа, изготовление которых требовало куда большего мастерства. Это событие приблизительно совпадает во времени с периодом быстрого увеличения мозга у эректусов, так что причинная связь между двумя событиями кажется вполне вероятной. Правда, что здесь было причиной, а что следствием, сказать трудно: можно привести доводы в пользу обеих точек зрения, но все — чисто умозрительные. Главное отличие ашельских орудий состоит в том, что им целенаправленно придавали определенную форму. Основным изделием стал нуклеус, причем его заостряли по периметру, так что получалось обоюдоострое ручное рубило, или бифас. Иногда рубила делались не из природного куска камня, а из предварительно отколотых кремневых отщепов. Вопрос о том, насколько резким или постепенным был переход от олдувая к ашелю, остается спорным. С одной стороны, известен так называемый развитой олдувай, трактуемый некоторыми экспертами как переходная культура, однако другие авторы оспаривают его переходный статус (Semaw et al., 2009). К позднеашельской эпохе (примерно 0,5 млн лет назад) форма орудий становится весьма совершенной и стандартизованной. Историк Б. Ф. Поршнев полагал, что эректусы делали свои орудия бессознательно, инстинктивно. В свете того, что мы знаем сегодня о поведении приматов, эту гипотезу можно смело отвергнуть. Даже умение колоть орехи камнями у шимпанзе не может стать инстинктивным — оно сохраняется в ряду поколений как культурная традиция, и каждой обезьяне приходится долго учиться этому искусству, подражая старшим. Несомненно, мастерство изготовления каменных орудий — и олдувайских, и ашельских — тоже было культурной традицией, знанием, передававшимся путем обучения и подражания. На слабость связей между африканскими и евразийскими популяциями эректусов указывает тот факт, что ашельские технологии долго не покидали пределов Африки. Многие популяции евразийских эректусов и их потомков продолжали пользоваться примитивными олдувайскими орудиями спустя сотни тысяч, даже миллион лет после того, как в Африке были изобретены бифасы. Например, галечные орудия олдувайского типа найдены на стоянках Бильцингслебен в Германии (400—300 тыс. лет назад) и Вертешсёллёш в Венгрии (300-200 тыс. лет назад). Есть аналогичные находки и в России (стоянка Карама в Горном Алтае). Лишь 500—350 тыс. лет назад позднеашельские орудия проникли через Ближний Восток на просторы Евразии (Деревянко, 2005). Скорее всего, это был не просто культурный обмен, а новая волна расселения выходцев из Африки. Причем это были уже не эректусы, а их более продвинутые потомки, которых сейчас классифицируют как "гейдельбергских людей (Homo heidelbergensis) в широком смысле". К сожалению, археологи находят каменные орудия на много порядков чаще, чем кости их производителей (камни лучше сохраняются), поэтому часто бывает трудно понять, кто какие орудия изготавливал. Но можно заметить, что время позднеашельской экспансии совпадает со временем разделения популяций предков сапиенсов и неандертальцев, которое определили недавно на основе анализа неандертальского генома (см. главу "Другое человечество"). Может быть, люди, принесшие позднеашельские технологии с Ближнего Востока в Европу, как раз и были представителями той предковой, еще не разделившейся популяции, от которой впоследствии произошли в Европе неандертальцы, а в Африке сапиенсы? Впрочем, если и так, то они, скорее всего, смешивались и с другими популяциями, то есть не были единственными предками двух "высших" видов людей. Первые европейцыВ раннем палеолите (ранний палеолит, он же нижний палеолит или ранний древнекаменный век, — период, охватывающий почти всю историю человечества от начала олдувайской культуры, зародившейся 2,6 млн лет назад, и вплоть до конца позднеашельской, которая начала сменяться среднепалеолитическими технологиями где-то около 0,2 млн лет назад) Европа никак не могла претендовать на статус центра цивилизации. Она была скорее задворками Ойкумены, "медвежьим углом". Но все же и сюда добирались отголоски прогресса, ковавшегося в те времена в основном в Африке. Древнейшим бесспорным свидетельством присутствия человека в Европе на сегодняшний день являются, по-видимому, человеческие кости, обнаруженные недавно в пещере Сима-дель-Элефанте ("Слоновая яма") в районе Атапуэрка на севере Испании. В 2008 году большая группа исследователей из Испании и США сообщила в журнала Nature о найденных здесь 32 примитивных каменных орудиях и части нижней челюсти человека с несколькими сохранившимися зубами (Carbonell et al., 2008). Самое замечательное в находке — это, конечно, ее возраст: 1,1—1,2 млн лет. Для определения возраста ученые сумели применить три независимых метода: биостратиграфический, палеомагнитный и радиометрический (О методах геохронологии см.: Марков А. В. Хронология далекого прошлого. http://elementy.ru/lib/430055). Высота пещеры Сима-дель-Элефанте составляет около 16 м, и в настоящее время она заполнена грунтом до потолка. Глина, песок и обломки камней заносились в пещеру с дождевыми потоками в течение нескольких сотен тысячелетий, пока не забили ее целиком. В пещерных отложениях выделено 13 последовательных слоев. Следы присутствия человека обнаружены в третьем слое снизу, примерно в четырех метрах от каменного дна пещеры. В одном слое с человеческой челюстью найдены кости мелких млекопитающих, в том числе мышевидных грызунов и хищников из семейства куньих. Среди этих зверьков есть виды, вымершие более миллиона лет назад. Значительно выше — на высоте и метров, между слоями 10 и 11 — удалось провести надежную магнитостратиграфическую границу, возраст которой составляет 0,78 млн лет. Человеческая челюсть, следовательно, намного древнее указанной даты. Кроме того, удалось использовать сравнительно малоизвестный радиометрический метод, основанный на распаде "космогенных" радиоактивных изотопов 26Аl и 10Ве. Эти изотопы образуются с постоянной скоростью под воздействием космических лучей в кристаллах кварца, находящихся на поверхности земли. Если камень попадает в пещеру, изотопы 26Аl и 10Ве перестают образовываться и только распадаются. По содержанию этих изотопов можно определить время, проведенное данным кусочком кварца в защищенном от космических лучей месте. При помощи этого метода ученые определили возраст двух образцов, один из которых был взят на 40 см выше человеческой челюсти, а другой на 4 м ниже, у самого дна пещеры. Возраст захоронения этих камней оказался равен 1,22 ± 0,16 и 1,13 ± 0,18 млн лет. На основании всего комплекса данных авторы заключили, что наиболее вероятный возраст человеческих останков из Сима-дель-Элефанте — от 1,1 до 1,2 млн лет. Вместе с человеческой челюстью найдены примитивные каменные орудия олдувайского типа, а также кости крупных млекопитающих из семейства полорогих, расколотые, чтобы извлечь мозг, и несущие характерные царапины от каменных орудий. До сих пор древнейшими ископаемыми остатками людей на территории Европы считались кости Homo antecessor возрастом около 0,8 млн лет, найденные той же бригадой ученых в том же районе Северной Испании в 1995 году (Carbonell et al., 1995). Правда, в других районах Южной Европы найдены каменные орудия, которые, возможно, являются еще более древними, однако возраст этих археологических находок не удалось определить с той же степенью точности, как находку из Сима-дель-Элефанте. По единственному обломку нижней челюсти трудно определить видовую принадлежность находки. По некоторым признакам эта челюсть напоминает эректусов и даже более примитивных людей из Дманиси. Другие признаки, напротив, являются более продвинутыми. В целом, по мнению авторов, лучше всего челюсть соответствует характеристикам вида Homo antecessor; описанного ими же в 1995 году. Впрочем, назвать этот вид "одной из популяций поздних, продвинутых эректусов" тоже не будет большой ошибкой. Не под запретом и версия "ранние Homo heidelbergensis". Ученые предполагают, что поздние эректусы, скорее всего, пришли в Западную Европу с востока. К тому времени, когда в Испании появились первые люди (1,1—1,2 млн лет назад), эректусы, по-видимому, уже были широко распространены в южных районах Азии. Кто-то из них мигрировал в Западную Европу, и здесь, на дальних задворках тогдашней Ойкумены, дал начало новому виду (расе, популяции?) людей — Н. antecessor. Череп Н. antecessor демонстрирует необычную смесь черт неандертальцев и сапиенсов. Крупные надбровные дуги, длинная и низкая черепная коробка, массивная нижняя челюсть без подбородка и крупные зубы сближают его с неандертальцами. Лицо, напротив, было относительно плоским и не выдавалось вперед, что сближает антецессоров с сапиенсами. Объем мозга около 1000 см3 — скорее как у эректусов и гораздо ниже средних значений для сапиенсов и неандертальцев. Ископаемые остатки животных, найденные вместе с костями Н. antecessor, говорят о том, что эти люди были умелыми охотниками на крупных зверей. Кроме того, они, несомненно, были каннибалами. Человеческие тела разделывались теми же приемами, а кости так же раскалывались для извлечения мозга, что и кости диких животных. Те и другие свалены древними обитателями пещеры в Атапуэрке в одну мусорную кучу. Это древнейшие бесспорные свидетельства каннибализма у людей. Детальный анализ находок привел исследователей к выводу, что каннибализм у Н. antecessor, по-видимому, не был связан с какими-либо ритуалами. Людей свежевали и разделывали теми же методами, что и другую добычу. Это не было и крайним средством выживания в голодные времена. Каннибализм был обыденностью и служил целям чисто гастрономическим. Среди съеденных были и дети, и взрослые. Вероятно, группы антецессоров конкурировали друг с другом и при случае с удовольствием пополняли свой рацион мясом соседей. Такое поведение иногда встречается и у шимпанзе (см. раздел "Обезьяньи войны" в главе "Эволюция альтруизма", кн. 2). Может быть, антецессоры впоследствии внесли вклад (наряду с носителями позднеашельских технологий?) в становление европейских популяций гейдельбергского человека, а те, в свою очередь, дали начало неандертальцам. Homo antecessor, судя по его морфологии, мог бы претендовать и на роль предка Н. sapiens, однако необходимо помнить, что первый вид жил в Европе, а второй родом из Африки. Вперед, на север. Недавно выяснилось, что и север Европы был заселен людьми гораздо раньше, чем принято было считать. Об этом свидетельствует находка, сделанная археологами в Пейкфилде (Восточная Англия) в 2005 году. В речных отложениях возрастом около 700 тыс. лет обнаружено 32 обработанных кремня. Орудия уже не олдувайские, а скорее ашельские, значительно более совершенные (Parfitt et al., 2005). Это самые древние следы присутствия человека в Северной Европе. В то время Британия соединялась с материком, а климат там был мягче, чем сейчас. В одном слое с орудиями обнаружены кости гиппопотама, льва, гигантского большеротого оленя Megaloceros dawkinsi, вымершего слона Palaeoloxodon antiquus, ископаемые остатки многих других млекопитающих, насекомых и растений. Обильный палеонтологический материал вкупе с рядом других независимых источников данных (стратиграфических, палеомагнитных) позволил достаточно надежно определить возраст пейкфилдской находки. Помимо прочего, авторы применили аминокислотный метод определения возраста, основанный на том, что "левые" аминокислоты, из которых состоят белки всех живых организмов, стечением времени рацемизируются, т.е. превращаются в смесь "правых" и "левых" форм. Находка показала, что древнее европейское человечество обладало высоким расселительным потенциалом. Все ранее известные следы присутствия человека к северу от Альп имеют возраст не более 500 тыс. лет. Пейкфилдские орудия, очевидно, были изготовлены поздними Н. antecessor или ранними Н. heidelbergensis (многие антропологи, впрочем, сомневаются в целесообразности разделения древнейших европейцев на два вида). Жизнь вокруг очагаГешер Бенот Яаков — знаменитая ашельская стоянка на территории Израиля в северной части рифта Мертвого моря. Люди ("гейдельбергские в широком смысле") жили там 750— 790 тыс. лет назад. Это была болотистая местность на берегу древнего озера. Уже многие десятилетия там работают археологи, добывая все новые сведения о жизни людей в те далекие времена. Превосходная сохранность органического материала позволила сделать выводы об ассортименте питания наших предков: кроме мясной пищи они употребляли разнообразные растения, включая орехи, зерна злаков, оливы, фрукты (среди прочих остатков ученые даже нашли изюминки). Древние обитатели этих мест пользовались различными орудиями труда — бифасами, отщепами, палками, некоторые из которых были отшлифованы (это древнейшие шлифованные деревянные предметы, известные на сегодняшний день). Кроме того, уже тогда люди пользовались огнем и умели поддерживать его в течение долгого времени. Об этом можно судить по обожженным предметам, найденным на стоянке. Не все предметы несут следы огня, а только их малая часть — значит, предметы обрабатывались огнем намеренно, а не вследствие пожаров (Goren-Inbar et al., 2004).
Предки современных людей, жившие 700—800 тыс. лет назад в долине Мертвого моря, разделывали слоновьи головы с помощью деревянных кольев и каменных орудий. По материалам сайта Института археологии Еврейского университета в Иерусалиме. http://www.archaeology.huji.ac.il. Результаты исследований дают неожиданно объемную картину жизни древних людей. Было обследовано более 80 тыс. микро- и макроартефактов и естественных остатков — орудия труда из кремня, известняка и базальта, их осколки, деревянные орудия, кусочки коры, древесины, скорлупа орехов, желуди, семена растений, раковины моллюсков, панцири крабов, остатки водных и наземных позвоночных. В одном из культурных слоев имеются свидетельства быстрого и внезапного захоронения. Это значит, что ископаемые остатки сохранили приблизительно такое расположение, какое им придали древние обитатели прибрежной стоянки. Жилое пространство стоянки неоднородно: оно делится на две части. В этих частях — юго-восточной, где располагался очаг (древнейший бесспорный очаг на сегодняшний день), и северо-западной — находок очень много. А между ними на пространстве в 5—6 метров плотность находок заметно ниже. Ассортимент предметов в этих частях неодинаков. Около очага концентрируются базальтовые и известняковые орудия труда — рубила, отщепы, скребки, наковальни и "молотки" для колки орехов. Кремневых орудий и кремневых осколков немного. Вокруг очага довольно много и древесных остатков, но, как ни удивительно, среди них почти нет кусочков со следами огня (таких найдено всего два). Также весьма впечатляет набор съедобных растений, среди которых нашлись водяные орехи, колючая кувшинка (Euryale ferox), желуди, оливы, лесной виноград (Vitis sylvestris), а также стиракс лекарственный (Styrax officinalis). За этими растениями приходилось ходить в лес, до которого от стоянки был неблизкий путь. Скорлупа орехов и желудей обожжена — значит, прежде чем расколоть орехи, люди обрабатывали их огнем, так орехи легче трескаются. Вокруг очага ученые нашли и остатки нескольких крупных крабов. В северо-западной части стоянки концентрируются кремневые отщепы, большей частью необожженные, и базальтовые осколки. А еще там много рыбьих костей. Добычей древних рыболовов были в основном крупные усачи-барбусы (до 1 м). Сравнение с естественным разнообразием рыб древнего озера говорит то ли о привередливости древних рыболовов, то ли об умении ловить лишь определенные виды рыб. Кроме того, их добычей были и черепахи. Останков млекопитающих довольно много: слоны, представители собачьих и парнокопытных и, естественно, вездесущие грызуны. Но они распределены по площади более или менее равномерно. Ясно, что около огня предки людей кололи орехи, ели крабов, использовали каменные орудия. А отойдя от костра на некоторое расстояние, они могли заняться изготовлением орудий, раскалывали камни, ели рыбу и — иногда — припасенные орехи. Зато обглоданные кости бросали повсюду (Alperson-Afil et al., 2009). Таким образом, функциональное разграничение жилья возникло еще в нижнем палеолите, задолго до появления "высших" представителей рода человеческого, сапиенсов и неандертальцев (древние обитатели Гешер Бенот Яаков вполне могли иметь отношение к предкам обоих видов). В каждой части жилого пространства принято было выполнять определенные действия. Очаг служил средоточием жизни социума, а подальше от него нужно было делать менее разнообразную, "черновую" работу. Едва ли такая сложная и организованная жизнь была бы возможна без членораздельной речи или хотя бы какого-то ее прообраза. "Анатомически современные люди"По-видимому, примерно от 800 до 300—200 тыс. лет назад большая и разношерстная совокупность полуразобщенных человеческих популяций, условно объединяемых под общим ярлыком Homo heidelbergensis, развивалась на просторах Африки, Европы, юго-западной и центральной Азии. Средний объем мозга этих людей был почти таким же, как у нас с неандертальцами (до 1100—1400 см3). С.В. Дробышевский (2010) отмечает, что особенно сильно у гейдельбергских людей развилась теменная доля, в первую очередь ее надкраевая область. Любопытно, что надкраевая извилина теменной доли — это один из участков мозга, который возбуждается у современных экспертов, овладевших палеолитическими технологиями, когда они изготавливают ашельские рубила, но не когда они мастерят более примитивные олдувайские орудия (см. главу "Душевная механика", кн. 2). Форма мозга Н. heidelbergensis свидетельствует о резком прогрессе в области контроля за движениями, в том числе способностей к прогнозированию и планированию своих будущих действий. Необходимо также отметить бурное развитие области, обеспечивающей согласование речи и движений рук, а также рельефное выступание зоны Брока, свидетельствующее о начале использования речи (Дробышевский, 2010).
Гейдельбергские люди (Homo heidelbergensis). Африка, Европа, Азия, 0,8—0,13 млн лет назад. Гейдельбергские люди были умелыми охотниками, о чем свидетельствуют, в частности, метательные копья возрастом 400 тыс. лет, найденные возле Шёнингена (Германия) в 1995 году (Thieme, 1997). Копья сделаны из стволов молодых елей, центр тяжести у них расположен так же, как у современных метательных копий, острые концы обожжены для твердости. Примерно 300—200 тыс. лет назад в некоторых из этих популяций начался плавный переход от позднеашельских технологий (относимых еще к нижнему палеолиту) к более сложным — среднепалеолитическим. В среднем палеолите двусторонние ручные рубила — бифасы, характерные для ашеля, — по-прежнему оставались в моде, но к ним добавилось много нового. Главным отличием среднепалеолитической каменной индустрии является широкое распространение орудий, изготовленных из отщепов, предварительно отколотых от "ядрища". В частности, вошло в моду изобретенное несколько раньше "леваллуазское расщепление" — весьма хитрый способ производства плоских каменных орудий с острыми краями (Древнейшие образцы леваллуазских изделий найдены на нижнепалеолитической (ашельской) стоянке Гешер Бенот Яаков, о которой говорилось выше). Камень сначала аккуратно обтесывали по краям, делая выпуклую заготовку, напоминающую черепаху (следы сколов похожи на пластины черепашьего панциря). Затем сбоку готовили "ударную площадку" и, наконец, точным ударом по этой площадке "черепаху" откалывали от основного камня. Получалось почти (или совсем) готовое плоское лезвие с одной гладкой стороной и одной — "черепаховой". Кроме новых методов изготовления орудий из специально подготовленных ядрищ и отщепов к числу важных среднепалеолитических новшеств относятся составные орудия — копья с каменными наконечниками (хотя достоверных находок таких орудий в среднем палеолите немного). В это время широко распространилось использование огня, а заодно и обычай готовить на нем пищу. Смена технологий имела место и в Африке, и в Европе, и в Азии, хотя в разных регионах были свои особенности. В Европе переход от раннего к среднему палеолиту примерно совпадает с важным эволюционным событием: на смену гейдельбергским людям пришли их потомки — неандертальцы. О них мы поговорим подробнее в главе "Другое человечество". В Африке приблизительно в это же время появляются первые люди, анатомически почти неотличимые от нас с вами. Их относят к виду Homo sapiens, но чаще называют осторожно "анатомически современными людьми". Тем самым подчеркивается то обстоятельство, что, хотя их анатомия была уже почти совсем как у нас, по своей культуре они еще до нас "не дотягивали". В частности, не было украшений, признаков наличия искусства и религиозных обрядов. Все это появилось несколько позже. Каменных дел мастера По-видимому, для того чтобы в совершенстве овладеть леваллуазской технологией — одним из важнейших "ноу-хау" среднего палеолита, — требовались годы упорного труда. Почти каждый современный специалист по палеолиту в общих чертах представляет, как делались леваллуазские лезвия, но лишь единицы в состоянии изготовить их сами, и эти единицы долго тренировались. Дело это непростое, требующее самого настоящего мастерства, что само по себе уже заставляет задуматься, так ли уж сильно уступали нам люди среднего палеолита в умственном развитии. По мнению антрополога Томаса Винна и психолога Фредерика Кулиджа из университета Колорадо, люди, способные овладеть этой технологией (придумали ее гейдельбержцы, но в совершенстве овладели неандертальцы и сапиенсы), должны были обладать совершенно определенными интеллектуальными способностями. У них, вероятно, была уже вполне современная по своей мощности долговременная рабочая память (ДРП) (Wynn, Coolidge, 2004). Так называют ту часть памяти, на которую полагаются эксперты в своей профессиональной деятельности, изумляя окружающих своим "невероятным" мастерством. В ДРП хранятся сложные блоки информации о методах решения конкретных задач. В состав этих блоков входят как "процедурные", так и "декларативные" элементы, то есть как моторные (двигательные) навыки и умения, так и понимание причинно-следственных связей в рамках данного круга задач, что позволяет успешно решать их в самых разнообразных условиях. Для извлечения нужного блока и перевода его в кратковременную рабочую память (КРП; о ней мы поговорим подробнее в главе "В поисках душевной грани") используются так называемые поисковые ключи и поисковые структуры (retrieval structures). Например, когда опытный шахматист ведет вслепую сеанс одновременной игры на 20 досках, он не запоминает по отдельности положение каждой фигуры на каждой доске. Он использует выработанные с годами тренировок "ключи", или "структуры", примерно такие: "доска №2 — сицилианская защита". На самом деле, конечно, экспертные поисковые структуры значительно более сложны, и для их выработки действительно требуются годы тренировок. КРП локализуется в лобных долях и отвечает за удержание внимания на тех идеях и блоках информации, с которыми сознание работает в данный момент. Некоторые авторы предполагают (и мне эта идея кажется правдоподобной), что "последний эволюционный штрих", сделавший разум уже совершенно таким, какой он у нас сейчас, состоял в небольшом увеличении объема КРП. Это не только позволило совершать более сложные действия, требующие одновременного внимания к нескольким объектам или идеям, но и открыло дополнительную свободу для мысленных экспериментов, новаторства и творчества. Но стать настоящим мастером в каком-то ремесле — будь то изготовление леваллуазских орудий или ковка по железу — можно и без "расширенной" КРП. Главное, что для этого нужно, — могучая и надежная ДРП, где хранятся необходимые мастеру навыки. Как и положено долговременной памяти, ДРП не локализована в каком-то одном отделе мозга, а рассеяна по многим участкам коры (почему это так, мы обсудим в главе "Душевная механика", кн. 2). Высочайшее мастерство среднепалеолитических каменных дел мастеров в сочетании с медленным техническим прогрессом и редкостью инноваций хорошо согласуется с идеей о том, что с ДРП у них уже все было в порядке, а вот с КРП — еще не очень. Впрочем, в таких рассуждениях всегда необходимо помнить, что умными нас делает не мозг сам по себе, а мозг в сочетании с культурой, то есть с теми знаниями, которые в него записываются в ходе общения с родителями и прочими соплеменниками. Можно иметь совершенно такой же, как у нас, сапиентный мозг, но, попав в неблагоприятные для сохранения культуры условия, деградировать чуть ли не до нижнепалеолитического уровня — аборигены Тасмании тому пример (см. ниже). Я почти уверен, что можно было бы и эректуса с мозгом в 900 кубиков приобщить к современной культуре, так что он мало отличался бы по умственному развитию от нас, сапиенсов постиндустриальной эпохи. Конечно, если взять на воспитание ребенка, а не взрослого с уже сложившимися стереотипами и чувством собственной важности. О гейдельбержцах с неандертальцами и говорить нечего. Объем КРП и у современных людей сильно варьирует. Ну были бы эти неандертальцы в среднем чуть менее блистательными новаторами, чем мы. Среди нас тоже блистательных новаторов не густо. Но ведь и культура не с неба падает. Вполне возможно, что в течение среднего палеолита ситуация висела на волоске. В какой из человеческих популяций начнется культурный перелом, известный нам как верхнепалеолитическая революция, могло зависеть от множества случайностей, в том числе культурно-социальных и демографических. Может, и не было никаких различий в объеме КРП между сапиенсами и неандертальцами. Может, уже у общих предков неандертальцев и сапиенсов мозг был по своему интеллектуальному потенциалу абсолютно таким же, как у нас сегодняшних. Не хватало только культуры. Возможно, мы в какой-то момент начали обгонять неандертальцев в развитии просто потому, что нам "повезло": мы жили в Африке, нас было больше, а численность и плотность населения способствуют культурному прогрессу. А может, у нас действительно развилась чуть более эффективная КРП, и это повысило вероятность того, что мы их обгоним, а не они нас. Или сначала мы получили крошечное преимущество из-за "африканской прописки" и плотности населения, наше поведение немного усложнилось, а уже потом благодаря эффекту Болдуина это повлекло за собой небольшое расширение КРП. Как узнать правильный ответ — трудно даже вообразить. Но в науке уже не раз так бывало, что кто-то торжественно объявит о принципиальной неразрешимости той или иной научной проблемы, а пройдет несколько лет — и все замечательно разрешается. Именно такая история произошла с химическим составом Солнца (кто бы мог подумать, что люди научатся определять состав звезд по их спектру!). Так что не будем отчаиваться. Мы еще вернемся к вопросу о сравнении интеллектуального потенциала сапиенсов и неандертальцев в главе "Другое человечество". История открытия древнейших сапиенсов была довольно драматичной. Уникальные находки были сделаны в районе поселка Кибиш на берегу реки Омо в Южной Эфиопии. Это местонахождение было обнаружено в 1967 году экспедицией Кенийских национальных музеев под руководством Ричарда Лики, которому тогда было всего 23 года. Тогда же были сделаны и главные находки — два человеческих черепа, названные Омо I и Омо II. Посланный на разведку в труднодоступный в то время район своим отцом, знаменитым палеоантропологом Луисом Лики, двадцатитрехлетний Ричард получил исчерпывающее представление об "африканской экзотике" — достаточно сказать, что при переправе через реку Омо экспедиция едва не досталась на обед крокодилам. Выкопанные сотрудниками экспедиции у селения Кибиш человеческие кости привели юного исследователя в полный восторг. Каково же было его разочарование, когда прибывшие на место раскопок родители — Луис и Мэри Лики (та самая, что описала следы австралопитеков в Лаэтоли) — вместо похвал и поздравлений устроили ему нагоняй, заявив, что он совершенно напрасно тратит деньги с их гранта на откапывание "анатомически современных" людей. По их мнению, ему следовало сосредоточиться на поисках более древних гоминид — австралопитеков, хабилисов и им подобных. С годами значение находки было переосмыслено. В сентябре 2008 года вышел специальный выпуск журнала Journal of Human Evolution, посвященный результатам их многолетнего изучения. Пожалуй, самое главное достижение состоит в уточнении датировок. Возраст черепов Омо I и Омо II долго оставался спорным. Это было связано прежде всего с несовершенством тогдашних методов радиометрического датирования. Результат первых радиоуглеродных датировок был — "старше 40 тыс. лет". Это означало просто-напросто, что кости слишком древние для радиоуглеродного анализа. По соотношению изотопов 230Th/234U в раковинах нильских устриц был определен возраст 130 тыс. лет (раковины были найдены чуть выше человеческих костей), но эта датировка признавалась ненадежной даже ее авторами. Привлечение дополнительных данных, в том числе биостратиграфических (по сопутствующим остаткам животных и растений) тоже не помогало прояснить ситуацию. Так продолжалось вплоть до самого конца прошлого века. Наконец в 1999 году большая команда американских антропологов решила взяться за дело всерьез. В течение четырех сезонов (1999, 2001, 2002, 2003) исследователи проводили в районе Кибиш широкомасштабные полевые работы. На анализ собранных материалов ушло еще несколько лет. Стратиграфия формации Кибиш теперь разработана с величайшей подробностью и точностью. Вся изученная толща делится на четыре части (пачки), причем человеческие кости происходят из первой, самой нижней. Отложения эти образовались в результате разливов реки Омо, полноводность которой менялась циклически с периодом около 25 тыс. лет (каждая пачка соответствует одному циклу). Цикличность связана с колебаниями климата, которые, в свою очередь, были обусловлены регулярными изменениями наклона земной оси. Такие же циклические колебания величины разливов были характерны и для Нила. Ученым удалось скоррелировать осадочные толщи в устье Нила (о них мы говорили в предыдущей главе) с соответствующими слоями формации Кибиш, и это стало одной из основ для новой уточненной датировки черепов Омо I и Омо II. Другим важным достижением стало датирование двух прослоев вулканического пепла, один из которых расположен непосредственно под костеносным слоем, а другой — значительно выше. Возраст нижнего прослоя, определенный по соотношению изотопов аргона, оказался равным 196 ± 2 тыс. лет, верхнего — 104 ± i тыс. лет. Вся совокупность данных свидетельствует о том, что наиболее вероятный возраст обоих черепов — 195 тыс. лет, причем величина возможной ошибки не превышает 5 тыс. лет. Это означает, что человеческие кости из формации Кибиш являются самыми древними костными остатками "анатомически современного человека", известными на сегодняшний день. Ранее основное внимание уделялось черепам, хотя, кроме них, было выкопано также несколько фрагментов посткраниального скелета Омо I. В 1999—2003 годах было найдено много новых костей, в том числе фаланги пальцев и часть бедра того же индивидуума. Скрупулезное изучение всех этих костей подтвердило, что Омо I, Омо II и их сородичи были "анатомически современными" людьми, то есть представителями вида Homo sapiens, но с отдельными архаичными чертами, которые сближают их с неандертальцами (Не все эксперты разделяют точку зрения о "сапиентности" черепов Омо I и Омо II. По мнению С. В. Дробышевского, у них все-таки слишком много архаичных признаков, чтобы относить их к виду Homo sapiens. Тем не менее в дальнейшем изложении мы будем придерживаться более общепринятой трактовки этих находок как "анатомически современных людей"). Важно, что такие же архаичные признаки имеются и у некоторых других древних сапиенсов, в том числе у доисторических обитателей пещер Схул и Кафзех в Израиле (к ним мы еще вернемся). Эти "неандертальские" признаки трактуются некоторыми исследователями как возможное свидетельство межвидовой гибридизации между сапиенсами и неандертальцами. В свете новых данных приходится признать, что некоторые из этих архаичных признаков могли быть унаследованы вышедшими из Африки древними сапиенсами от своих африканских предков, то есть от еще более древних сапиенсов. В этом случае предположение о смешанных браках с неандертальцами вроде бы становится излишним. С другой стороны, это предположение подтверждается новейшими данными палеогенетики, о чем будет сказано ниже. Сапиенсы все-таки скрещивались с неандертальцами, причем, по-видимому, как раз в то время (порядка 120—80 тыс. лет назад) и в том районе (Ближний Восток), где жили древние обитатели пещер Схул и Кафзех. Вместе с человеческими костями в формации Кибиш найдены многочисленные скелетные остатки млекопитающих, птиц и рыб. Среди них практически нет вымерших видов: все эти животные и поныне обитают в Восточной Африке. Природная среда в этом районе 200 тыс. лет назад была примерно такой же, как сейчас, только климат был несколько более влажным, а местность — более болотистой. К слову, заметим, что Африка — единственный континент, где деятельность первобытных охотников, по-видимому, не привела к значительному сокращению разнообразия крупных животных. Значительно большее негативное влияние оказали наши предки на фауну Евразии, где они, вероятно, ускорили вымирание мамонтовой фауны, а в Австралии и обеих Америках приход человека и вовсе привел к катастрофическим последствиям (см. главу "Великое расселение сапиенсов"). Почему древние сапиенсы никого не истребили только на своем родном континенте? Может быть, потому, что африканская фауна эволюционировала вместе с гоминидами в течение нескольких миллионов лет и африканские экосистемы имели достаточно времени, чтобы приспособиться к повадкам двуногих хищников? Важным результатом полевых работ 1999—2003 годов стала обширная коллекция каменных орудий из нижних слоев формации Кибиш (ранее там были найдены лишь единичные орудия). Это более или менее типичная индустрия африканского среднего палеолита (Африканский средний палеолит (в отличие от европейского) по непостижимым для неспециалистов причинам (связанным с хронологией и типами орудий) принято называть не средним палеолитом, а "средним каменным веком" (middle stone age). Это чревато чудовищной путаницей, потому что существует ведь еще и мезолит, который тоже переводится как "среднекаменный век", но это совершенно другая эпоха, которая наступила после палеолита (и до которой мы в этой книге даже не доберемся). Из сострадания к читателям и самому себе я буду называть африканский "средний каменный век" средним палеолитом) с большой долей бифасов (обоюдоострых орудий), без каких-либо верхнепалеолитических изысков вроде костяных иголок или тонко обработанных лезвий и уж подавно — без украшений и произведений искусства. Таким образом, нет оснований утверждать, что древнейшие сапиенсы с берегов реки Омо по технологическому уровню хоть в чем-то превосходили своих современников — ранних европейских неандертальцев, изготавливавших примерно такие же среднепалеолитические каменные орудия. Впрочем, исследователи пока воздерживаются от каких-либо прямых утверждений об интеллектуальном и культурном уровне древнейших сапиенсов. Они отмечают, что сам вопрос о том, были ли эти люди "отсталыми" или "прогрессивными", неявно предполагает сравнение с верхним палеолитом Европы, который традиционно считается чем-то вроде "культурного эталона" для ранних представителей нашего вида (см. главу "Великое расселение сапиенсов"). Но почему выбран именно такой эталон? Исключительно в силу исторических причин: археология палеолита стала развиваться в Европе гораздо раньше, чем в других частях света. Бесспорно, люди, обитавшие в Эфиопии 200 тыс. лет назад, не были верхнепалеолитическими европейцами, ну и что с того? Исследователи саркастически замечают, что мы и Сократа сочли бы "отсталым" по сравнению с самым посредственным американским школьником, если бы в качестве мерила "прогрессивности" использовалось присутствие в соответствующем археологическом слое плееров и пластиковых бутылок. К сожалению, имеющихся данных пока недостаточно, чтобы судить о том, как жили и о чем думали древнейшие африканские сапиенсы (Journal of Human Evolution, 2008). Митохондриальная Ева и игрек-хромосомный Адам в африканском ЭдемеСравнительный анализ митохондриальной ДНК (мтДНК) и Y-хромосом современных людей показал, что все современное человечество происходит от небольшой популяции, жившей в Восточной Африке 160—200 тыс. лет назад (Сапп et al., 1987). Это подозрительно хорошо совпадает с археологическими данными: древнейшие "анатомически современные люди" найдены как раз в этом районе в отложениях именно такого возраста. Затем в какой-то момент, скорее всего между 100 и 60 тыс. лет назад (опять же, судя по результатам анализа мтДНК), небольшая группа сапиенсов вышла из Африки, и потомки этой группы впоследствии заселили весь мир: от них произошло все современное внеафриканское человечество (см. главу "Великое расселение сапиенсов"). С этим сегодня почти никто не спорит. Главный вопрос в том, только ли от них оно происходит. Смешивались ли африканцы по мере своего расселения с местными евразийскими популяциями или просто вытеснили их? В течение почти двух десятилетий (с конца 1980-х до второй половины 2000-х) перевес был на стороне второй точки зрения. Главным аргументом были упомянутые результаты анализа мтДНК и Y-хромосомы. Но как на основе анализа ДНК можно судить об истории и миграциях древних людей? Пожалуй, нам нужно поближе познакомиться с методикой и логикой таких исследований. Сначала на основе полиморфизма (разнообразия) нуклеотидных последовательностей мтДНК людей из разных регионов мира было реконструировано эволюционное (генеалогическое) дерево этой части человеческого генома. Для таких реконструкций существуют специально разработанные, весьма надежные методики. Это похоже на то, как лингвисты восстанавливают историю различных списков с одного исходного текста по изменениям (заменам, пропускам и добавлениям букв или слов), которые неизбежно накапливаются от переписчика к переписчику. Например, если в одном списке имеются ошибки А и Б, в другом — А, Б и В, в третьем — только Б, в четвертом — Б и Г, то логичнее всего предположить, что самый древний список — третий. С него были сделаны первый и четвертый, а затем с первого был сделан второй. Когда тексты длинные (и ошибок поэтому достаточно много), результаты получаются вполне достоверные. Если известно, в каких географических точках были найдены соответствующие рукописи, то можно восстановить пути миграций соответствующих копий. В случае с ДНК вместо ошибок переписчиков используются случайные мутации (в основном нейтральные, то есть не влияющие на приспособленность и потому незаметные для отбора). Это даже удобнее, потому что люди — и, соответственно, их ДНК — размножаются с определенной частотой, более регулярно, чем средневековые переписчики копировали ту или иную рукопись. К тому же типичная частота возникновения мутаций приблизительно известна. Это аналогично тому, как если бы каждая рукопись переписывалась, например, один раз в 20 лет, а все переписчики делали примерно одинаковое количество ошибок на каждую тысячу слов. Тогда по количеству различий между двумя рукописями можно было бы определить время существования их "общего предка". Для молекул ДНК это вполне возможно (степень точности — вопрос отдельный). Оказалось, что если построить эволюционное дерево мтДНК современных людей и двигаться по его ветвям сверху вниз (из настоящего в прошлое), то все ветви в итоге сходятся в одну точку во времени и пространстве: Восточная Африка, 160—200 тыс. лет назад. Так появилась в научной печати и в СМИ "митохондриальная Ева" (митохондрии, как мы помним, передаются по материнской линии), а вслед за ней аналогичным образом возник и "Y-хромосомный Адам" (Y-хромосома есть только у мужчин и передается от отца к сыну), живший примерно в то же время и в том же месте. Эти результаты были восприняты общественностью очень бурно, и, как водится, мало кто понял их истинный смысл. На самом деле ничего удивительного нет ни в Адаме, ни в Еве. Любые гомологичные (то есть имеющие общее происхождение) участки ДНК где-нибудь в прошлом неизбежно сходятся в одну точку, то есть в одну предковую молекулу ДНК. И эта точка вовсе не обязательно совпадает с моментом возникновения вида. Более того, если брать разные гомологичные участки ДНК, каждый из них даст свою, отличную от других "точку схождения". Откуда берутся митохондриальные Евы Любая популяция любого вида животных обязательно имела в прошлом свою митохондриальную Еву — последнюю общую праматерь всех ныне живущих представителей данной популяции по прямой материнской линии. Евы появляются автоматически и неизбежно из-за случайных колебаний частот генетических вариантов (например, вариантов митохондриальной ДНК) в популяции. Чтобы понять, почему так получается, рассмотрим простую модель. Допустим, у нас есть популяция, включающая десять самок, у каждой из которых имеется свой вариант мтДНК, немного отличающийся от остальных. Число самок (то есть размер популяции) не имеет принципиального значения: оно влияет только на среднее число поколений, требующихся для того, чтобы одна из этих самок стала Евой. Чем больше популяция, тем дольше придется ждать. Предположим, что каждая самка оставляет после себя с равной вероятностью либо ноль дочерей, либо одну, либо две. Обозначим исходные варианты мтДНК у наших десяти самок буквами латинского алфавита: abcdefghij (1) Это поколение 1. Чтобы смоделировать следующее (второе) поколение, воспользуемся генератором случайных чисел. Сойдет и обычная игральная кость. Нам нужна последовательность из десяти случайных целых чисел в диапазоне от нуля до двух, чтобы определить, сколько дочерей родила каждая самка. Бросаем кость десять раз, если выпадает один или два, записываем ноль, если три или четыре — записываем единицу, если пять или шесть — двойку. У меня получилась такая последовательность: 2212210011 В соответствии с этими числами "родим" для каждой самки дочерей. Учитывая, что дочь наследует мтДНК матери, выпишем распределение митохондриальных гаплотипов (вариантов) в поколении 2: aabbcddeefij (2) Можно заметить, что из десяти исходных гаплотипов во второе поколение перешли только восемь. Две самки (g и h) не оставили дочерей, и их митохондриальные линии пресеклись. Самок у нас теперь стало 12, поэтому для того, чтобы смоделировать поколение 3, понадобится 12 случайных чисел. Вот они: 220020212021 А вот и третье поколение: aaaaccddeeeiij (3) В третьем поколении "потерялись" еще два гаплотипа: b и f. Продолжая моделирование, получаем последовательность поколений: aaaeeejj (4) eeejj (5) ejjjj (6) eejjjjjj (7) eeejjj (8) eeejj (9) eeeej (10) eeeejj (11) eeeeeej (12) eeeejj (13) eeeej (14) eeeej (15) eeeej (16) eeeeee (17) Вот и все: к семнадцатому поколению в нашей популяции остался только один митохондриальный гаплотип из десяти исходных. Это значит, что прямые потомки по женской линии остались только у одной из исходных десяти самок. Процесс этот абсолютно неизбежен: сколько бы мы ни взяли исходных самок с разными гаплотипами, через какое-то число поколений в популяции останется только один из них. При этом одна из древних носительниц этого гаплотипа автоматически превращается в митохондриальную Еву — последнюю общую праматерь всех особей в популяции по непрерывной женской линии. Кстати, какая именно самка в нашей модели стала Евой для поколения №17? Думаете, это самка "е" из поколения 1? А вот и нет: у поколения 17 есть и более поздняя общая праматерь. Это самка с гаплотипом "е" из поколения 6. Самка "е" из поколения 1, конечно, тоже является общей праматерью поколения 17 по женской линии, но она не самая поздняя из таких праматерей. При желании можно придумать фантастические ситуации, в которых Ева никогда не появится. Но эти ситуации не имеют отношения к реальности. Например, Евы не будет, если каждая самка непременно оставляет после себя хотя бы одну дочь (нет ни бездетных самок, ни таких, кто оставил после себя лишь сыновей). Каждому ясно, что так не бывает. Обязательно какая-то часть самок умирает, не родив ни одной дочери. Исходное множество гаплотипов может не сократиться до одного и в том случае, если численность популяции будет бесконечно расти. Но так тоже не бывает: планета не резиновая, рост любой популяции рано или поздно останавливается.
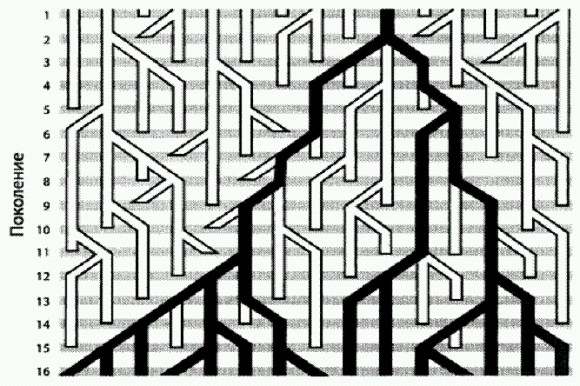
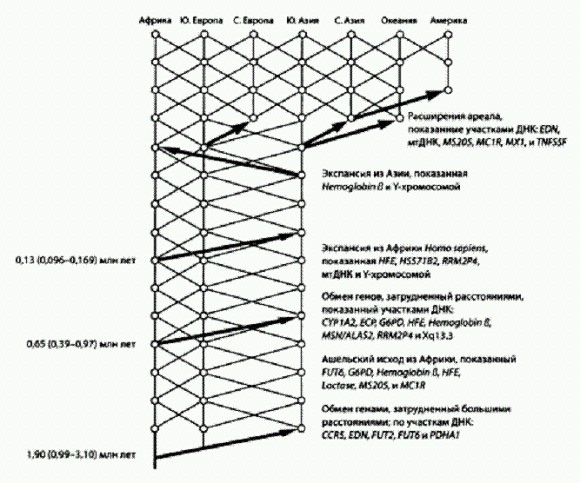
Вымышленный пример генеалогического древа вариантов митохондриальной ДНК. В каждом поколении в популяции присутствует 15 самок, у каждой из которых может быть ноль, одна, две или три дочери. Все самки 16-го поколения восходят к митохондриальной Еве из поколения №2. Что будет, если популяция разделится на две — например, часть особей переселится на другой материк, и каждая из двух дочерних популяций будет процветать на своем материке? В этом случае в каждой из двух популяций тоже обязательно рано или поздно закрепится какой-то один из исходного набора гаплотипов. Причем, скорее всего, на разных материках это будут разные гаплотипы. Получится, что у каждой из двух популяций есть своя митохондриальная Ева. Кроме того, будет и третья Ева — общая для обеих групп. Она окажется глубже в прошлом, чем обе "персональные" Евы разделившихся популяций. По мнению ряда экспертов, приблизительное совпадение результатов по мтДНК и Y-хромосоме — не более чем случайность, отчасти объясняющаяся тем, что оба этих участка генома имеют общее свойство: они присутствуют у каждого человека лишь в одном экземпляре (точнее, в одном варианте: "экземпляров" гораздо больше, они есть в каждой клетке, но все одинаковые). Большинство других участков генома — любые участки ядерных неполовых хромосом — присутствуют в двух вариантах, один из которых получен от отца, другой от матери. Есть еще Х-хромосома, занимающая промежуточное положение: у женщин она присутствует в двух экземплярах, у мужчин в одном. Американский антрополог и генетик Алан Темплтон еще в 2005 году обратил внимание на тот факт, что ожидаемое время схождения эволюционного дерева, построенного для отдельного участка ДНК, в одну точку зависит от того, в скольких вариантах присутствует данный участок в организме (Templeton, 2005). Быстрее всего должны сходиться как раз мтДНК и Y-хромосома (что и наблюдается, они сходятся 160— 200 тыс. лет назад). Это не значит, что именно тогда и появился Н. sapiens. По мнению Темплтона, это значит лишь, что данные участки генома не годятся для реконструкции более давних событий. Участки, локализованные на Х-хромосоме, сходятся в более далеком прошлом (до 2 млн лет); все остальные участки — в еще более глубокой древности, некоторые даже еще до того, как разделились эволюционные линии человека и шимпанзе. Как это может быть? Очень просто: если бы мы могли проследить во всех деталях родословную предков человечества — не какого-то конкретного человека, а всей популяции вместе, вплоть до общих предков с шимпанзе, — мы увидели бы, что эта родословная никогда не "схлопывалась" до одного-единственного индивида. Всегда, в любой момент прошлого, существовала некая предковая группа особей с различающимися генами. Поэтому "наша" эволюционная линия вполне могла унаследовать от общих с шимпанзе предков не один, а сразу несколько вариантов того или иного гена. Впоследствии мы могли растерять этот исходный полиморфизм, так что у нас в итоге остались гены, происходящие лишь от одного из исходных вариантов. Именно так и произошло с мтДНК и многими другими генами. Но это не обязательно: по некоторым генам часть исходного полиморфизма могла досуществовать (и досуществовала) до наших дней. В общем, получается, что история мтДНК — еще не история человечества. Каким образом по мтДНК или другому участку генома можно сделать вывод о выходе наших предков из Африки в какое-то определенное время? Это возможно в том случае, если вскоре после данного события у кого-то из переселенцев возникла мутация в изучаемом участке ДНК, которая затем в ходе экспансии размножилась. И тогда современный генетик увидит, что частота встречаемости данной мутации у внеафриканского населения, к примеру, 10 %, а в Африке ее нет. Время возникновения мутации определяется на основе других, позднее возникших мутаций, по методу "молекулярных часов". Ну а если вскоре после выхода из Африки в данном участке генома никакой мутации не возникло? Тогда, разумеется, ничего не выйдет: этот участок генома просто не сохранит следов интересующей нас экспансии. Поэтому по одному-единственному участку генома (например, по мтДНК) нельзя делать окончательные выводы об эволюции и истории расселения человечества. Для таких выводов необходим комплексный анализ многих разных участков генома. Темплтон проанализировал помимо мтДНК и Y-хромосомы еще 23 участка генома и пришел к следующим выводам. Разные участки ДНК сохранили следы разных событий в истории человечества. Общая картина довольно точно совпадает с той, которая реконструируется по данным археологии. Три участка ДНК сохранили следы древнейшей волны выхода из Африки около 1,9 млн лет назад. Это означает, что в наших жилах течет кровь древних азиатских (а не только африканских) эректусов!
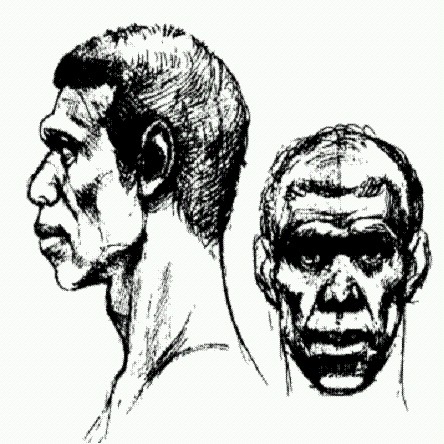
Схема истории человечества по А. Темплтону. Справа перечислены участки ДНК (гаплотипы), подтверждающие то или иное событие. Видно, что мтДНК и У-хромосома "работают" лишь начиная с последнего выхода из Африки около 100 тыс. лет назад. По рисунку из Templeton, 2005. Семь участков ДНК свидетельствуют о втором исходе из Африки около 0,65 млн лет назад. Это, наверное, позднеашельская экспансия. Гейдельбергские люди, представители этой волны, — тоже наши предки. Наконец, еще пять участков ДНК (в том числе мтДНК и Y-хромосома) подтверждают третий исход из Африки около 130 тыс. лет назад. Кроме того, данные Темплтона показывают, что обмен генами между евразийскими и африканскими популяциями наших предков практически никогда не прекращался, хоть и был сильно затруднен большими расстояниями. Получается, что древнее человечество все-таки не было совокупностью полностью изолированных популяций (рас, подвидов, видов?) — оно было относительно единым на протяжении двух последних миллионов лет. Выводы Темплтона оспаривались одними экспертами, в то время как другие разными путями приходили к похожим результатам. Но вплоть до публикации сенсационных результатов прочтения геномов неандертальца и денисовца (речь о которых пойдет ниже), преобладала точка зрения о полном вытеснении африканскими сапиенсами всех древних евразийских популяций. Три дырявые ракушки и начало "подлинно-человеческой" культурыВажнейшим переломным рубежом в истории Homo sapiens традиционно считается так называемая верхнепалеолитическая революция — качественный скачок в культурном и технологическом развитии, произошедший около 45—35 тыс. лет назад и примерно совпадающий по времени с вторжением сапиенсов в неандертальскую Европу. Еще сравнительно недавно ученые полагали, что именно в это время наши предки впервые научились изготавливать сложные изделия из кости и рога, освоили новые методы обработки камня (требующие не только высочайшего мастерства, но и способности концентрировать внимание на нескольких вещах одновременно), стали пользоваться украшениями и создали первые произведения искусства — наскальные изображения животных. Но люди современного типа, как мы уже знаем, появились в Африке значительно раньше — около 200 тыс. лет назад. Их культура поначалу была среднепалеолитической, то есть примерно такой же, как у других современных им человеческих рас, в том числе европейских неандертальцев. Около 130 тыс. лет назад сапиенсы появляются в Западной Азии, но эта первая волна "исхода из Африки", по-видимому, оказалась неудачной. Затем началась вторая волна расселения. Сапиенсы двигались на восток вдоль берегов Индийского океана, оставляя за собой характерные раковинные кучи, свидетельствующие о пристрастии к морепродуктам (подробнее см. в главе "Великое расселение сапиенсов"). Все современное неафриканское человечество — потомки этой второй волны переселенцев (с примесью генов местных евразийских популяций). До тех пор пока материальная культура сапиенсов оставалась среднепалеолитической, они не показывались в Европе, где господствовала более крепкая физически и хорошо приспособленная к холодному климату неандертальская раса. Верхнепалеолитическая революция дала сапиенсам возможность быстро (за 5—6 тыс. лет) оккупировать Европу. Некоторые находки последних лет, однако, несколько размыли эту относительно стройную картину. Выяснилось, что отдельные элементы верхнепалеолитической культуры были выработаны нашими предками задолго до вторжения в Европу, а возможно, даже до выхода из Африки. Древнейшие признаки зарождения "подлинно человеческой" духовной культуры выглядят довольно скромно. Это три продырявленные ракушки моллюска Nassarius gibbosulus, обнаруженные в 2006 году (Vanbaeren et al., 2006). Две из них, найденные в коллекции Британского музея естественной истории, происходят из пещеры Схул вблизи Хайфы (Израиль). В этом районе селились попеременно то неандертальцы (до 130 (Впрочем, датировки, указывающие на столь раннее присутствие неандертальцев в Леванте, спорны, и есть мнение, что какое-то время неандертальцы и сапиенсы могли проживать на этой территории совместно (Вишняцкий, 2010)) (65—47 тыс. лет назад), то сапиенсы (130—80 и после 47 тыс. лет назад). Анализ породы, прилипшей к одной из раковин, позволил установить, что раковина происходит из так называемого слоя В, возраст которого оценивается в 130— 100 тыс. лет. В это время здесь жили сапиенсы. Это были представители "первой волны" выходцев из Африки, о которой говорилось выше. В слое В пещеры Схул найдено десять скелетов, большинство из которых по своей анатомии соответствуют человеку современного типа с рядом архаичных черт (но есть и вполне неандерталоидные особи). Специалисты допускают, что по крайней мере некоторые из этих людей были сознательно похоронены соплеменниками. На грудь одного из покойников (Схул-5) сородичи положили нижнюю челюсть крупного кабана, что трактуется как свидетельство существования погребальных обрядов и религиозных верований.
Человек из пещеры Схул (рисунок, основанный на черепе Схул-5; возраст около 100 тыс. лет). Похожие отверстия в раковинах Nassarius gibbosulus после смерти моллюска могут возникать и естественным путем, но происходит это сравнительно редко (такие перфорации наблюдаются всего у 3,5 % современных раковин, собранных в море или на берегу). Таким образом, вероятность случайного попадания в пещеру сразу двух перфорированных раковин, при том что других раковин этого вида в пещере не обнаружено, составляет примерно одну тысячную. Попытки археологов самостоятельно проделать в раковинах Nassarius gibbosulus подобные отверстия при помощи среднепалеолитических каменных орудий увенчались полным успехом. Сначала надо кремневым острием пробить маленькую дырочку, а затем вращательными движениями придать отверстию нужную форму и размер. Раковины не могли попасть в пещеру без помощи человека, поскольку пещера находится на высоте 65 м над уровнем моря. 130-100 тыс. лет назад она тоже находилась достаточно высоко (не менее 45 м). Звери и птицы никогда не заносят этих моллюсков на такую высоту. Едва ли древние обитатели пещеры Схул могли принести моллюсков в свое жилище только для того, чтобы их съесть, потому что раковины эти мелкие (около полутора сантиметров) и пищевой ценности не представляют. Возможно, это все-таки остатки древнейших украшений. Третья перфорированная ракушка, относящаяся к тому же виду моллюсков, найдена в коллекции парижского Музея человека и происходит из среднепалеолитической стоянки на севере Алжира, в 150 км от моря. Точно определить возраст этой стоянки пока не удалось, но, по косвенным данным, он может быть сопоставим с возрастом слоя В пещеры Схул. Эти находки наряду с другими ставят под сомнение теорию взрывного, внезапного появления "подлинно человеческой" культуры на рубеже среднего и верхнего палеолита. По-видимому, культурное развитие наших предков протекало более постепенно, чем считалось до сих пор. Другой важный вывод, который можно сделать из этих открытий, состоит в том, что наши прямые предки — архаические Homo sapiens, — возможно, начали опережать неандертальцев по культурному уровню задолго до конца среднего палеолита. Обнаруженные сих пор свидетельства символического осмысления мира и ритуальной деятельности неандертальцев (захоронения, украшения) либо значительно моложе израильских и алжирских находок, либо их возраст вызывает сомнения, да и принадлежность их неандертальцам в ряде случаев не бесспорна. Дырявых ракушек становится больше, но не надолгоВ период от 80 до 60 тыс. лет назад произошло сразу несколько событий, сыгравших важную роль в развитии человечества. В это время происходили крупные миграции Homo sapiens в Африке, тогда же человечество распространилось за пределы родного континента и заселило юг Азии. В тот же период наши предки прошли через "бутылочное горлышко" — период резкого сокращения численности (может быть, их осталось всего около 15 тыс.). Это было связано, вероятно, с чудовищным извержением вулкана Тоба на Суматре, которое произошло 74 тыс. лет назад. Это была катастрофа не в пример нынешним: весь полуостров Индостан был засыпан толстым слоем вулканического пепла, атмосфера в течение нескольких лет едва пропускала солнечные лучи, что привело к сильному похолоданию (эффект ядерной зимы). И все же сапиенсы выжили, причем не только на своей родине в Африке, но, возможно, и в Индии, куда они уже успели к тому времени добраться (подробнее см. в главе "Великое расселение сапиенсов"). Вскоре после извержения Тобы произошли важные события в культурной эволюции африканских сапиенсов, в их "духовном развитии". В это время впервые в большом количестве появляются украшения (не три несчастные продырявленные ракушки, а целые ожерелья) и геометрические узоры, выцарапанные на камнях и скорлупе страусиных яиц. Многие антропологи считают, что появление украшений и орнаментов было связано с развитием символического мышления и речи. Древние ожерелья составлялись не как попало, а из ракушек определенного размера и оттенка. Возможно, они играли роль символов, рассказывающих о статусе человека и его роде-племени. Это в свою очередь предполагает существование большого, сложно структурированного общества, в котором представители разных общин имели схожие интересы и понятия и, видимо, могли эти интересы друг с другом обсуждать. Если не считать трех ракушек, о которых рассказано выше, украшения впервые по-настоящему вошли в обиход в Южной Африке, в двух археологических культурах, известных как стилбейская (СБ) и ховьесонс пурт (ХП). Культура СБ древнее, чем ХП. Для нее характерны тонко обработанные, заостренные с двух концов наконечники копий, костяные остроконечники, исцарапанные геометрическими узорами кости и камни, многочисленные ожерелья из ракушек. Для более молодой культуры ХП характерен несколько иной набор каменных и костяных артефактов, в том числе изделия с притуплёнными краями, которые, вероятно, были частями каких-то составных орудий, а также предполагаемые древнейшие костяные наконечники для стрел (хотя луков найти не удалось). Чтобы понять последовательность событий, происходивших на заре человеческой цивилизации, крайне важно получить точные датировки культур СБ и ХП. Разумеется, такие попытки предпринимались и раньше, причем довольно успешные, однако полной ясности до сих пор не было. Дело в том, что разные находки датировались разными людьми, в разных лабораториях и разными методами. Неудивительно, что общая картина получалась несколько смазанной. В частности, до сих пор никто не мог сказать наверняка, был ли между культурами СБ и ХП перерыв или культура СБ непосредственно переросла в культуру ХП; не ясна была и длительность периодов культурного расцвета. Кроме того, оставалось неясным, связаны ли вспышки культурного развития с изменениями климата; одновременно ли развивались культуры СБ и ХП в разных климатических зонах. На все эти вопросы попыталась ответить группа исследователей из Южной Африки, Австралии, Великобритании и Германии (Jacobs et al., 2008). Они получили максимально точные оптико-люминесцентные датировки для 44 проб грунта из археологических слоев, содержащих артефакты культур СБ и ХП, а также из непосредственно прилегающих к ним выше- и нижележащих слоев. Пробы были взяты в девяти районах археологических раскопок в Южной Африке, Лесото и Намибии. Таким образом, авторы получили 44 датировки, каждая со своими доверительными интервалами, которые затем были подвергнуты сложному статистическому анализу. Выяснилось, что культура СБ появилась примерно 71900 лет назад, просуществовала очень недолго — менее тысячелетия — и исчезла около 71 тыс. лет назад. Культура ХП появилась после долгого перерыва около 64800 лет назад, просуществовала примерно 5300 лет и исчезла около 59500 лет назад. Нового периода культурного расцвета африканским сапиенсам пришлось ждать уже до самой верхнепалеолитической революции, которая началась то ли в Европе, то ли в Азии и докатилась до Африки около 40 тыс. лет назад. Полученные даты хорошо согласуются с большинством датировок, полученных ранее другими методами. Однако только теперь стало ясно, что между двумя культурами был долгий перерыв. Украшения, изощренные орудия и другие признаки высокого культурного развития не сразу стали неотъемлемой чертой человеческих обществ: они появлялись, затем забывались, снова входили в обиход и снова исчезали. Авторам не удалось обнаружить никакой корреляции между культурным развитием древних обитателей Южной Африки и природными условиями. Обе культуры были широко распространены на юге Африки, причем они не были приурочены к каким-то определенным природным зонам. Люди, принадлежавшие к культурам СБ и ХП, жили и у берегов океана, и в глубине материка, и в горах, в районах с разным климатом, разной сезонностью дождливых и засушливых периодов. Похоже на то, что носители новой прогрессивной культуры быстро расселялись по большим территориям и одинаково успешно осваивали разные природно-климатические зоны. Причины взлета и упадка культур СБ и ХП пока не ясны. Некоторые авторы предполагают, что это могло быть связано с колебаниями плотности населения и интенсивности культурного обмена между поселениями. Возможно, для того, чтобы культурный прогресс стартовал, необходима некоторая пороговая численность популяции. Этот порог может быть ниже, если соседние поселения разнородны в культурном отношении и между ними существует интенсивный обмен знаниями. Чем меньше и разреженнее население и чем меньше культурных контактов между отдельными сообществами, тем выше вероятность забывания полезных знаний, утраты технологий и деградации культуры. Эти предположения основаны на результатах математического моделирования (Powell et al., 2009), и в них, на мой взгляд, содержится ключ к пониманию крупномасштабных закономерностей эволюции культуры. Возможно, именно этим механизмом объясняется удивительная и трагичная судьба аборигенов Тасмании. Около 8 тыс. лет назад, когда в Северном полушарии растаяли ледники, уровень океана поднялся, и Тасмания оказалась отрезанной от Австралии, с которой прежде составляла единое целое. Тасманийцы оказались в полной изоляции. В течение последующих тысячелетий их культура катастрофически деградировала. От уровня, характерного для австралийских аборигенов того времени (приблизительно соответствующего верхнему палеолиту), тасманийцы быстро "скатились" чуть ли не до нижнепалеолитического состояния. Они разучились делать костяные орудия, теплую одежду, каменные орудия с деревянными рукоятками, сети, рыболовные гарпуны, копьеметалки и бумеранги. Все это, по археологическим данным, было у них 8000 лет назад, но полностью исчезло к моменту появления европейцев (Нenrich, 2004). Мы вернемся к истории людей современного типа в главе "Великое расселение сапиенсов", после того как познакомимся с некоторыми данными генетики, а также с другими популяциями среднепалеолитических людей, живших в Европе и Азии одновременно с нашими прямыми предками - африканцами. |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Вверх |
||||
|
|
||||