|
||||
|
|
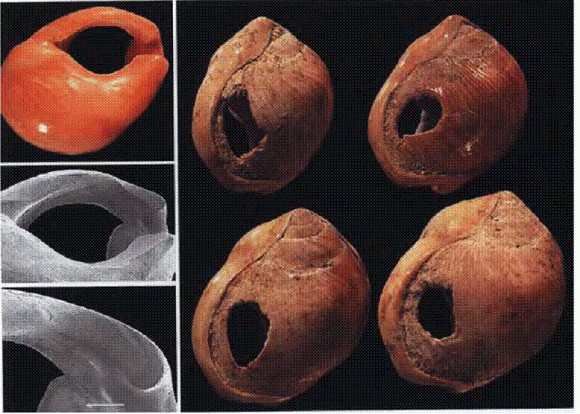
ГЛАВА 2. ОЧЕЛОВЕЧИВАНИЕОбезьяна берет каменный ножУ австралопитеков, с которыми мы познакомились в предыдущей главе, мозг был не больше, чем у шимпанзе. Поэтому кажется естественным думать, что и по уровню интеллекта они не превосходили нынешних "нечеловеческих человекообразных". С другой стороны, размер мозга — не идеальный показатель ума. У современных людей положительная корреляция между интеллектом и объемом мозга хоть и существует, но не является абсолютно строгой. Есть гении с маленьким мозгом, есть глупцы с большим (см. ниже). К тому же у австралопитеков, по данным С. В. Дробышевского, была увеличена теменная доля по сравнению с шимпанзе и другими "нечеловеческими" гоминоидами. Это может указывать на улучшенную координацию движений рук и способность совершать более тонкие манипуляции. Точно ли мы уверены, что австралопитеки и другие ранние гоминиды третьего — четвертого миллионолетия до нашей эры (парантропы, кениантропы) не делали ничего такого, что оказалось бы не под силу шимпанзе и гориллам; что им не приходилось решать интеллектуальных задач, перед которыми современные обезьяны, возможно, спасовали бы? В первой главе мы упоминали примитивные костяные орудия — обломки мелких костей, при помощи которых парантропы, по-видимому, охотились на термитов. Замечательный факт, но шимпанзе тоже добывают термитов при помощи палочек двух типов — "пробойников" и "удочек", которые предварительно подвергаются обработке: обезьяны удаляют лишние сучки, листья, а у "удочек" размочаливают конец, то есть не просто пользуются готовым орудием, а изготавливают его. Лишь в 2010 году стали известны факты, прямо указывающие на то, что австралопитеки все же делали нечто, не укладывающееся в рамки нормального поведения современных обезьян, кроме человека. Это сенсационное открытие сделано большим коллективом палеоантропологов из США, Германии и Франции. Начиная с 1999 года авторы проводили систематические раскопки в районе Дикика в Эфиопии. Самая известная из их находок — "дочка Люси", с которой мы познакомились в предыдущей главе. В январе 2009 года неподалеку от того места, где нашли "дочку Люси", ученые обнаружили две кости с царапинами от каменных орудий — фрагмент ребра крупного копытного размером с корову и обломок бедренной кости зверя помельче, размером с козу. Детальное исследование находок при помощи сканирующего электронного микроскопа и рентгеновской спектроскопии подтвердило, что царапины были оставлены на костях после смерти животных, но до фоссилизации (окаменения). Отметины недвусмысленно свидетельствуют о том, что каменные орудия использовались для срезания и соскребания мяса, а также для раскалывания костей (чтобы добраться до костного мозга) (McPherron et al., 2010). Главное в этих находках — их чрезвычайно древний возраст, который удалось определить при помощи комплекса радиометрических и стратиграфических методов: от 3,39 до 3,42 млн лет. До сих пор древнейшими свидетельствами использования гоминидами каменных орудий считались находки возрастом 2,5—2,6 млн лет, сделанные в районе Гона в нескольких километрах к западу от Дикики (Semaw et al., 1997), а также в ряде других районов Эфиопии и Кении (см. ниже).
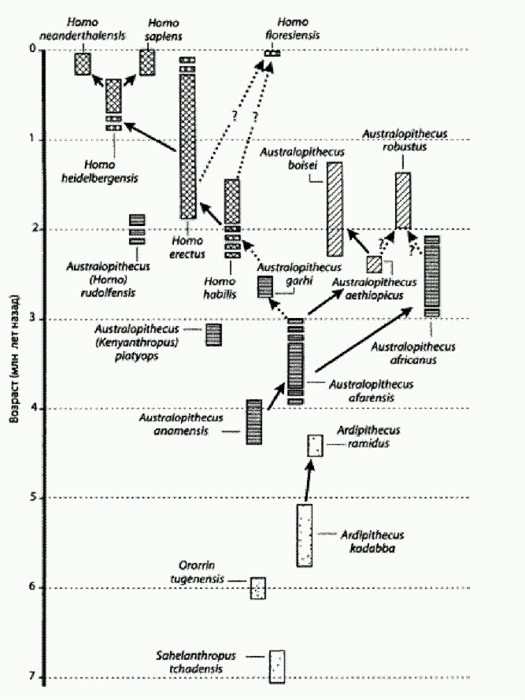
Austrolopithecus garhi. Эфиопия, 2,5 млн лет назад. Таким образом, новые данные удревнили начало использования каменных орудий — а также употребления в пищу мяса крупных травоядных — примерно на 800 тыс. лет. Долгое время считалось, что первооткрывателями каменной индустрии почти наверняка были ранние представители рода Homo, такие как Homo habilis (человек умелый) или Н. rudolfensis, а их предшественники — австралопитеки — каменных орудий делать не умели. Правда, древнейшие из известных орудий (около 2,6 млн лет назад) несколько старше самых старых костей, более или менее уверенно относимых к ранним Homo (около 2,3 млн лет назад). Однако это несоответствие списывали на то, что камни сохраняются гораздо лучше, чем кости. Новые находки сделали временной разрыв между первыми следами использования каменных орудий и древнейшими костными остатками хабилисов и рудольфензисов слишком большим, чтобы его можно было заполнять отговорками о редкой сохраняемости костей. Единственным представителем гоминид, обитавшим 3,4 млн лет назад в данном районе Африки, был австралопитек афарский — вид, к которому относится Люси, ее "дочка", Большой человек и многие другие неплохо сохранившиеся окаменелости. Таким образом, напрашивается вывод, что афарские австралопитеки научились использовать острые камни для разделки туш за миллион лет до появления хабилисов. К сожалению, ни орудий, ни каких-либо острых камней рядом с исцарапанными костями авторы не нашли. Поэтому вопрос о том, пользовались ли австралопитеки "готовыми" природными камнями с заостренным краем или сами изготавливали примитивные орудия, остается открытым. Кроме того, ясно, что если австралопитеки и умели делать примитивные каменные орудия уже 3,4 млн лет назад, то занимались они этим очень редко, иначе их изделия давно были бы найдены. Не исключено, что австралопитеки все-таки занялись всерьез изготовлением каменных орудий, но позже — 2,5—2,6 млн лет назад, незадолго до появления первых Homo. Каменные орудия этого возраста найдены в Эфиопии вместе с ископаемыми костями одного из поздних видов грацильных австралопитеков — Austrolopithecus garhi (этот вид был описан в 1997 году). Там же найдены кости травоядных животных, с которых этими орудиями сдирали мясо. Поскольку никаких следов присутствия других гоминид, кроме A. garhi, поблизости не наблюдается, логично предположить, что именно А. garhi и были изготовителями орудий. По объему мозга (около 450 см3) A. garhi не выделяется из общего ряда грацильных австралопитеков. Australopithecus sediba - австралопитек, похожий на человекаСерия замечательных открытий, сделанных палеоантропологами в последние десятилетия, показала, что эволюционная история африканских гоминид была сложной и многоплановой. Современные схемы эволюции гоминид напоминают скорее пышно ветвящийся куст, чем прямую и стройную линию, ведущую "от обезьяны к человеку". Многие признаки, характерные для Homo erectus и других продвинутых представителей рода людей, по-видимому, появлялись независимо в разных эволюционных линиях гоминид, стоявших на австралопитековом уровне развития (австралопитеки, парантропы, кениантроп) и на уровне ранних Homo (Н. habilis, Н. rudolfensis, а также ряд форм, известных по фрагментарному материалу и условно относимых к "ранним Homo"). Подобная мозаичность в распределении признаков и многочисленные параллелизмы при их формировании часто наблюдаются во время крупных эволюционных преобразований (см. главу "Направленность эволюции" в книге "Рождение сложности" (Марков, 2010)). Поэтому новые находки часто не столько проясняют, сколько усложняют картину. Первые Homo, несомненно, произошли от грацильных австралопитеков или близких к ним форм. Однако остается неясным, какой из видов австралопитеков был нашим прямым предком. На эту роль с переменным успехом претендуют Australopithecus africanus ("ребенок из Таунга"), A. afarensis (Люси с "дочкой"), A. garhi и Kenyanthropusplatyops. Конечно, сохраняется некоторая (не слишком большая) вероятность того, что прямой предок первых Homo еще не найден, а известные науке формы являются боковыми веточками — более или менее близкими родственниками искомого предка. Неясно также, кто именно из ранних Homo (Н. habilis, Н. rudolfensis или какой-то пока не открытый вид) дал начало человеку прямоходящему (Н. erectus), который уж точно был нашим предком (а также предком гейдельбергского человека и неандертальца). Морфологические различия между австралопитеками и ранними Homo не очень велики. О непреодолимой пропасти говорить явно не приходится. Граница между родами Australopithecus и Homo была проведена условно, методом "волевого решения". Эксперты договорились (с подачи знаменитого антрополога Луиса Лики, одного из первооткрывателей хабилисов), что все формы, у которых мозг больше б00 см3, будут считаться людьми, у кого меньше — австралопитеками. Но и это различие "поплыло", когда обнаружились особи Н. habilis с объемом мозга всего 510 см3. Это меньше, чем у некоторых парантропов. По мнению ряда антропологов, Н. habilis лучше было бы относить к роду Australopithecus. Другие предлагают "разбросать" образцы, ныне относимые к хабилисам, на несколько видов, кого-то включив в род людей, кого-то — в род австралопитеков. Это лишний раз показывает, насколько условны границы между родами и видами ископаемых гоминид. Но все-таки австралопитеков можно отличить от ранних Homo по некоторым деталям строения черепа, зубов и посткраниального скелета. Именно этот разрыв и заполнила новая находка, сделанная южноафриканскими палеоантропологами под руководством Ли Бергера (того самого, который доказал, что "ребенок из Таунга" погиб в когтях орла). Неполные скелеты двух представителей нового вида гоминид — мальчика и взрослой женщины — найдены в пещере Малапа в 15 км к северо-востоку от знаменитых местонахождений Стеркфонтейн и Сварткранс (Berger et al., 2010). Чуть позже там нашли еще и фрагменты костей рук маленького ребенка. Первую кость нового вида гоминид — правую ключицу мальчика-подростка — нашел 15 августа 2008 года девятилетний Мэтью, сын Ли Бергера. Отец включил Мэтью в список соавторов, но рецензенты журнала Science не одобрили эту идею. Возраст находки — от 1,95 до 1,78 млн лет. Эти цифры можно считать вполне надежными, потому что определение возраста проводилось при помощи трех разных методов: радиометрического уран-свинцового датирования (анализы проводились независимо в двух лабораториях — в Берне и Мельбурне), палеомагнитного и биостратиграфического (по сопутствующей фауне). Вместе со скелетами мальчика и женщины обнаружены кости двух видов саблезубых кошек (Dinofelis и Megantereon), лесного кота, бурой гиены, гиеновидной собаки, двух видов мангустов, лошади, зайца и пяти видов вымерших и доживших до наших дней антилоп. Все эти животные погибли, провалившись в естественную ловушку — узкий провал, ведущий в глубокую подземную полость. Попав в пещеру, их тела стали недоступными для хищников и падальщиков. Из верхней части пещеры частично разложившиеся тела были смыты вниз потоком, образовавшимся в результате сильных ливней. В яме на дне подземной реки останки были быстро погребены под слоем осадка. Мальчику было по человеческим меркам 12—13 лет (это определили по состоянию коренных зубов). Он был немного выше взрослой женщины и имел более развитую мускулатуру. Очевидно, мужчины этого вида были крупнее женщин, но ненамного. Половой диморфизм у этих гоминид был примерно таким же, как у современных людей. Авторы не без колебаний включили новооткрытый вид в род Australopithecus под видовым названием sediba, что означает "родник" на местном африканском языке сесото. Некоторые антропологи, в том числе Дональд Джохансон, первооткрыватель Люси, видевший кости A. sediba, выразили сомнения в правильности такого решения. По их мнению, эту гоминиду следовало отнести к роду Homo. Основная часть статьи Бергера и его коллег посвящена скрупулезному анализу признаков черепа, зубов и посткраниального скелета А. sediba. В статье приведена огромная, занимающая четыре с половиной страницы таблица признаков, по которым новый вид можно отличить от других видов австралопитеков и ранних людей. Из 130 рассмотренных признаков 23 отличают A. sediba от австралопитеков и сближают с людьми, семь, наоборот, сближают новый вид с австралопитеками и отличают от людей. Из всех австралопитеков A. sediba больше всего похож на А. africanus. В приведенной таблице есть пять признаков, по которым А. sediba сходен с A. africanus и ранними Homo, но отличается от других австралопитеков. Остальные признаки позволяют отличить A. sediba от некоторых австралопитеков и людей, но не дают дополнительных доводов в пользу отнесения его к тому или другому роду. Самым важным "австралопитековым" признаком A. sedibа является маленький объем мозга (420 см3 у мальчика; в ходе дальнейшего роста его мозг должен был увеличиться еще примерно на 5%). По строению рук и ног A. sediba тоже больше похож на австралопитеков, чем на людей. Однако в строении черепа, зубов и таза наблюдается много человеческих черт. К их числу относятся надглазничный валик, очень похожий на таковой у Н. habilis и Н. erectus, другие детали строения лобных костей, слабо выступающие скулы, характерная форма носовых костей, свидетельствующая о более выступающем (по сравнению с австралопитеками) носе, небольшие по размеру коренные зубы и ряд особенностей таза. По-видимому, A. sediba ходил на двух ногах еще быстрее и увереннее, чем его предки австралопитеки. Его походка, по мнению авторов, была почти такая же, как у эректусов, которые были превосходными ходоками. Те признаки в строении таза, которые отличают A. sediba от австралопитеков и сближают с Н. erectus, говорят о более эффективном распределении нагрузок при ходьбе и более развитых мышцах — разгибателях бедра. По-видимому, у A. sediba уже были почти человеческие ягодицы (у человекообразных обезьян эта часть тела развита гораздо слабее, чем у людей). Таким образом, находка подтвердила, что в эволюции гоминид полноценная человеческая походка сформировалась раньше, чем началось увеличение мозга. Предком A. sediba, скорее всего, был A. africanus. Авторы предполагают, что новый вид мог отделиться от своего предка значительно раньше, чем 2 млн лет назад. Найденные скелеты не могут принадлежать прямым предкам Homo, потому что древнейшие представители рода людей, найденные в Эфиопии и предположительно относящиеся к виду Н. habilis, имеют более почтенный возраст — 2,33 млн лет. Однако первые Homo могли произойти от более древних популяций А. sediba. Точно определить положение A. sediba на эволюционном дереве гоминид пока невозможно, для этого потребуются дополнительные находки. Не исключено, что ранние представители этого вида или их близкие родственники дали начало первым людям — Н. habilis или Н. rudolfensis, а может быть, даже напрямую Н. erectus, потому что несколько второстепенных признаков сближают A. sediba с эректусами "через голову" хабилисов и рудольфензисов. Распутать этот клубок трудно, потому что все формы, о которых идет речь, — и поздние австралопитеки, и ранние люди — приходились друг другу очень близкой родней (примерно как современные шимпанзе и бонобо, если не ближе). По-видимому, в период между 2,5 и 1,8 млн лет назад в Южной и Восточной Африке жили разнообразные популяции продвинутых гоминид, подвергавшиеся сходному действию отбора и эволюционировавшие более или менее в одном направлении, но с разной скоростью. Какие-то из этих популяций скрещивались друг с другом, какие-то нет. Подразделить эту разношерстную компанию на "еще обезьян" (австралопитеков) и "уже людей" (ранних Homo) можно лишь чисто условно. Никаких "объективных критериев" для такого разделения найти не удается. Мозг начинает расти. Зачем?К концу третьего миллионолетия до н. э. у некоторых представителей многочисленной и разнообразной группы африканских двуногих обезьян наметилась новая эволюционная тенденция — началось увеличение мозга. Первых гоминид, у которых мозг стал больше, чем у современного шимпанзе, традиционно относят к виду Homo habilis (человек умелый). Ископаемые остатки этого вида имеют возраст примерно от 2,3 до 1,5 млн лет. Многие антропологи выделяют часть "ранних Homo" с более крупными зубами, чем у типичных хабилисов, и уплощенным лицом в отдельный вид Homo rudolfensis. Рудольфензисы жили одновременно с хабилисами в тех же районах Восточной Африки, и вообще-то не исключено, что это были не две разных популяции, а одна и та же, просто ее представители отличались высокой вариабельностью. Хабилисы, по всей видимости, произошли от грацильных австралопитеков, таких как А. afarensis, A. garhi, A. africanus или A. sediba. Рудольфензисов некоторые авторы сближают с кениантропом, который жил 3,5 млн лет назад (одновременно с A. afarensis). Иногда кениантропа не выделяют в отдельный род, а рассматривают как одного из грацильных австралопитеков. Главный общий признак рудольфензисов и кениантропов — уплощенное лицо. По поводу многих находок, относящихся ко второй половине третьего миллионолетия, нет полной ясности, относить ли их к австралопитекам или ранним Homo. Впрочем, для нашего рассказа эти подробности не очень важны. Мы позволим себе впредь для простоты называть всю пеструю компанию ранних Homo просто хабилисами. По данным С. В. Дробышевского (2007, 2010), для мозга хабилисов по сравнению с австралопитеками характерно усиленное развитие участков, которые у современных людей связаны с речью и координацией движений рук. Таким образом, хабилисы — первые гоминиды, у которых заметны признаки эволюционного движения в сторону развития речи. Все, что происходило в эволюции гоминид до начала увеличения мозга — уменьшение клыков, совершенствование двуногого хождения, изменения стопы, предполагаемые изменения в социальной жизни, — можно рассматривать как чисто "обезьяньи дела". Если бы какие-то из этих гоминид, не изменившись, дожили до наших дней, — можете не сомневаться, они бы сейчас сидели в зоопарке. Ходили бы по вольеру на своих двоих, заводили моногамные семьи, жевали фрукты и бифштексы. И даже их маленькие, почти совсем человеческие клыки не вызывали бы у публики чувства общности с этими "животными". Пропуск в мир людей дает интеллект, а для интеллекта нужны мозги. О том, почему у хабилисов начал расти мозг, существует не меньше гипотез, чем о происхождении бипедализма. Мы еще вернемся к этой теме (см. главу "Общественный мозг", кн. 2). Но несколько первых, самых очевидных соображений нужно упомянуть сразу. Рост мозга при всех оговорках, исключениях и отступлениях от общего правила все-таки предполагает поумнение. Исходя из того, что известно на сегодняшний день о работе мозга, можно заключить, что простое увеличение числа нейронов в коре — даже без какой-то особой реструктуризации и прочих хитростей — может автоматически вести к росту интеллекта, то есть к способности решать более сложные задачи. Память "записывается" в межнейронных контактах (синапсах) (см. главу "Душевная механика", кн. 2). Больше нейронов — больше возможностей для формирования таких контактов — больше объем памяти. Больше информации в голове — больше возможностей принимать умные решения.
Kenyanthropus platyops. Кения, 3,5 млн назад. Пусть это рассуждение не является строгим научным доказательством, но все же едва ли рост мозга у ранних Homo мог иметь какой-то иной адаптивный смысл, кроме развития когнитивных (то есть умственных, познавательных) способностей. Анатоль Франс ничего не доказывает Положительная корреляция между размером мозга и интеллектом бесспорно существует. Она не является абсолютно строгой (коэффициент корреляции меньше единицы), но из этого вовсе не следует, что "размер не имеет значения". Корреляции такого рода никогда не бывают абсолютно строгими. Коэффициент корреляции всегда меньше единицы, какую бы зависимость мы ни взяли: между массой мышцы и ее силой, между длиной ног и скоростью ходьбы и так далее. Действительно, встречаются очень умные люди с небольшим мозгом и глупые — с крупным. Часто в этом контексте поминают Анатоля Франса, у которого объем мозга был всего 1017 см3 — нормальный объем для Homo erectus и гораздо ниже среднего для Homo sapiens. Это, однако, вовсе не противоречит тому, что интенсивный отбор на интеллект способствует увеличению мозга. Для такого эффекта вполне достаточно, чтобы увеличение мозга хоть немного повышало вероятность того, что особь окажется умнее. А вероятность, безусловно, повышается. Внимательно рассмотрев таблицы объема мозга великих людей, часто приводимые в качестве опровержения зависимости ума от размера мозга, нетрудно убедиться, что у подавляющего большинства гениев мозг все-таки крупнее среднего. Анатоль Франс и несколько других известных людей с маленьким мозгом на этом фоне выглядят самыми обычными статистическими "выбросами" (outliers), без которых не обходится никакая закономерность в биологии. Так, например, курение вызывает рак легких, невзирая на то, что кто-то курил всю жизнь и дожил до 90, а кто-то не курил и умер от рака легких.
Homo rudolfensis. Восточная Африка, 2,3—млн лет назад. По-видимому, нестрогость рассматриваемой корреляции во многом объясняется неизбежным элементом стохастики (случайности) в процессе роста мозга у эмбрионов и детенышей. Откуда берется стохастика в индивидуальном развитии — вопрос сложный и явно выходящий за рамки этой книги. Я ограничусь тем, что напомню читателю об одном очевидном факте. Мозг устроен гораздо сложнее (для его описания нужно больше бит информации), чем геном, который с некоторыми оговорками можно рассматривать как "программу" развития организма, в том числе и мозга. В геноме человека лишь 3 х 109 нуклеотидов, каждый из которых содержит два бита информации. Не будем мелочиться: для ровного счета пусть будет 1010 бит. Структура межнейронных связей мозга, от которой, собственно, и зависит его производительность, требует для своего описания как минимум в миллион раз большего носителя: в ней не менее 1016 бит информации (на самом деле, вероятно, на несколько порядков больше, см. главу "Душевная механика", кн. 2). Таким образом, в геноме физически невозможно записать структуру мозга во всех деталях (даже если речь идет о мозге новорожденного). В геноме закодированы лишь общие принципы формирования и роста нейронов. Все остальное является результатом процессов самоорганизации на манер тех, что приводят к образованию снежинок из маленьких и одинаковых молекул воды. Все снежинки сложные и красивые, и все чуть-чуть разные, хотя и сходные в общих чертах. Это пример неизбежной стохастики в процессах самоорганизации. При реализации генетической программы развития некоторый элемент стохастики неизбежен еще и потому, что любые две клетки хоть чуть-чуть, но отличаются друг от друга как по числу молекул ключевых белков, так и по поведению (второе является следствием первого). А если вспомнить о существовании наследственной изменчивости по множеству генов, не влияющих на размер мозга, но влияющих на тонкие детали функционирования нейронов (см. главу "Генетика души", кн. 2), то становится очевидно, что положительная корреляция между размером мозга и уровнем интеллекта никак не может быть абсолютной.
Homo habilis. Восточная Африка, 2,3-1,5 млн лет назад.
Размер мозга связан не только с интеллектом, но и с размером организма: крупному животному при прочих равных требуется более крупный мозг. Ученые, разумеется, учитывают это обстоятельство в своих расчетах. Например, во многих случаях вместо абсолютного объема мозга рассматривают вычисляемый по специальной формуле коэффициент энцефализации, отражающий соотношение объема мозга и размеров тела. Рост мозга у ранних Homo свидетельствует о том, что отбор стал благоприятствовать особям с более крупным мозгом. Иными словами, более умные особи стали иметь больший репродуктивный успех — оставлять в среднем больше потомков, чем их менее сообразительные сородичи. Учитывая социальность гоминид, можно предположить, что действие естественного отбора опосредовалось социумом: коллективы, в которых было больше умных индивидов, возможно, предоставляли более благоприятные условия для размножения своих членов, чем коллективы, состоявшие из недоумков. Выигрыш от ума наверняка был существенным, потому что мозг — орган дорогостоящий. Его рост кроме пользы приносит и очевидный вред. Иначе, надо полагать, многие животные давно стали бы мудрецами. Мозг потребляет много энергии — значит, животному нужно больше пищи. Большеголовых детенышей трудно рожать (повышается смертность при родах). Эти недостатки должны перевешиваться пользой от ума, иначе мутации, ведущие к увеличению мозга, будут отбраковываться отбором. Очевидно, предки ранних Homo столкнулись с некими новыми жизненными задачами, успешное решение которых, во-первых, приносило значительную пользу (повышало репродуктивный успех), во-вторых, требовало экстраординарных умственных усилий по сравнению с тем, что до сих пор привычно делали австралопитеки. Теперь, когда мы корректно сформулировали проблему, нужно просто посмотреть, что же изменилось в жизни ранних Homo или их ближайших предков по сравнению с более ранними австралопитеками — насколько об этом можно судить по имеющимся скудным данным. Два новшества определенно появились в их жизни. Во-первых, судя по археологическим находкам, с середины третьего миллионолетия до н. э. началось систематическое изготовление каменных орудий. Мы помним, что пользоваться острыми камнями, по-видимому, могли еще афарские австралопитеки 3,4 млн лет назад, но изготовление орудий началось много позже. Самые старые орудия примерно на 0,2—0,3 млн лет древнее самых старых костей хабилисов. Возможно, более древних хабилисов просто еще не нашли, но, как уже говорилось, может быть и так, что начали изготавливать орудия еще австралопитеки. Тем более что кости A. garhi найдены по соседству с древнейшими орудиями, а костей других гоминид там нет. В любом случае наши предки, по-видимому, сначала стали делать орудия, а потом у них начал расти мозг. Это абсолютно нормально. Сначала изобретается новая сложная манера поведения, сохраняющаяся в популяции как культурная традиция. Новый навык дается с трудом, на него едва хватает мозгов, ему учатся полжизни, зато в случае успеха он приносит большую пользу. Похожая ситуация наблюдается в тех популяциях шимпанзе, где существует традиция колки орехов при помощи молота и наковальни (см. главу "В поисках душевной грани", кн. 2). Если шимпанзе додумались колоть орехи, то и австралопитеки вполне могли додуматься раскалывать кости крупных травоядных, чтобы извлечь мозг. Может быть, с этого все и началось, а идея соскребать с костей остатки мяса острым каменным обломком появилась позже. Закрепление новой культурной традиции меняет характер отбора: теперь мутации, повышающие скорость обучения новому поведению или эффективность его осуществления, будут закрепляться. Культурное новшество меняет направленность эволюции. Между изменившимся поведением и закреплением облегчающих его морфологических адаптаций, таких как увеличенный мозг, вполне может быть заметный временной зазор. Подробнее об этом механизме, который называется эффектом Болдуина, мы поговорим в главе "Генетика души" (кн. 2). Древнейшая, так называемая олдувайская (См. сводную таблицу этапов развития палеолитической культуры в конце первого тома на с. 452), каменная индустрия была примитивна — нужно было просто найти два подходящих камня и бить один о другой, чтобы отколоть отщеп с острым краем. Форма изделия не имела значения — как отколется, так и ладно, лишь бы край получился острым. Но все же такая деятельность, по-видимому, лежит за пределами умственных способностей современных нечеловеческих обезьян. Например, антропологи несколько лет пытались научить бонобо Канзи — настоящего обезьяньего гения, который добился замечательных успехов в освоении человеческой речи (Зорина, Смирнова, 2006), — изготавливать каменные орудия, похожие на олдувайские. Несмотря на все усилия экспериментаторов, успехи Канзи в производстве орудий были довольно скромными. Он научился раскалывать камни, бросая один на другой сверху. При этом действительно получаются обломки с острыми краями, которыми можно пользоваться как орудиями. Но технология, применявшаяся хабилисами, — держать в одной руке "ядрище", в другой "молот" и откалывать отщепы точными ударами, — по-видимому, оказалась для Канзи слишком заумной. Он пытался так работать, подражая экспериментаторам, но удары получались слишком слабыми. Исследователям так и не удалось научить Канзи производить изделия, полностью схожие с олдувайскими (Schick et al., 1999). Итак, вот первая новая задача, которая могла стимулировать рост мозга у ранних Homo. Пресловутый тезис "труд сделал из обезьяны человека" имеет право на существование. Но этим дело не исчерпывается. Кроме появления каменных орудий было еще одно важное изменение, которое состояло в том, что ранние Homo, по-видимому, начали систематически включать в свой рацион мясо крупных животных. Каменные орудия они использовали для разделки туш и соскребания мяса с костей. Для того-то их и делали. Австралопитеки почти наверняка не охотились на крупных животных, а лишь иногда подбирали остатки трапез хищников. Вопрос о том, когда гоминиды из падалыциков превратились в охотников, крайне спорный. В 1950—1960-е годы Луис Лики на основе находок, сделанных им в Олдувайском ущелье на севере Танзании, отстаивал точку зрения, согласно которой древние люди (хабилисы, а затем и ранние эректусы) охотились на крупную дичь уже 1,8 млн лет назад. Впоследствии более тщательный анализ следов, оставленных на костях травоядных зубами хищников и каменными орудиями, привел многих экспертов к выводу, что олдувайские гоминиды все-таки добирались до туш травоядных после хищников, то есть были падалыциками. Об этом свидетельствует тот факт, что следы от каменных орудий на костях крупных травоядных идут поверх следов зубов крупных хищников. Это значит, что хищники первыми добирались до жертвы, а людям доставались объедки. Добыча падали в плейстоценовой африканской саванне вряд ли была похожа на увеселительную прогулку. Не стоит называть ее "сбором падали", как делают некоторые авторы. Это вам не сбор грибов и ягод. Конкуренция за мертвые туши крупных травоядных в те времена, скорее всего, была устрашающей. Особенно в конце сухого сезона: это самый голодный период в саванне, когда пригодной для гоминид растительной пищи почти не оставалось. Двуногие обезьяны должны были соперничать с опасными хищниками, включая саблезубых кошек и крупных гиен. Кроме того, им наверняка приходилось жестоко конкурировать с другими группами себе подобных. Нужны были хитрость, смелость, готовность идти на риск ради коллектива, нужна была четкая слаженность действий. Все это — новые задачи, которые тоже можно смело включить в список вероятных стимулов увеличения мозга. Средневековые падальщики Тезис о том, что добыча падали в саванне была для ранних Homo делом нервным и опасным, трудно проиллюстрировать конкретными живыми примерами. Среди современных охотников-собирателей такое поведение встречается редко, а главное, все они прекрасно умеют охотиться и в основном этим и занимаются. Да и падаль нынче не та, что в плейстоценовой африканской саванне. Довольно адекватное, как мне кажется, представление о психологической обстановке, которая складывается вокруг дармовой горы мяса — туши крупного животного, — когда за нее конкурируют несколько человеческих групп, можно получить из фрагмента древнеисландской "Саги о Греттире" (перевод О. А. Смирницкой): Весною подул с севера сильный ветер и дул почти неделю. Когда же он стих, люди пошли посмотреть, что им выбросило на берег. Торстейном звали человека, жившего на Мысе Дымов. Он нашел кита, прибитого к южному берегу мыса, в месте, что зовется Реберные Скалы. Это был большой синий кит. Торстейн сразу же послал человека в Залив, к Флоси, и на ближние хутора. Эйнаром звали человека, жившего на Дворе Расселина. Он жил на земле людей Холодной Спины и должен был смотреть за всем, что прибивало к берегу по ту сторону фьорда. Он заметил, что прибило кита, тут же взял лодку и стал грести через фьорд к Загонному Заливу. Оттуда он послал человека на Холодную Спину. Узнав про это, Торгрим с братьями спешно собрались, их было двенадцать человек на десятивесельной лодке. Поехали с ними и сыновья Кольбейна, Ивар с Лейвом, числом всего шестеро. Добравшись до места, все пошли к киту. Теперь надо рассказать про Флоси. Он послал за своими родичами на север, к Ингольвову Фьорду и Офейгову Фьорду, и за Олавом, сыном Эйвинда, который тогда жил у Столбов. Флоси и люди с Залива первыми подошли к киту. Они взялись разделывать тушу, а отрезанные куски оттаскивали на берег. Сперва их было человек двадцать, но скоро народу прибавилось. Тут подоспели на четырех лодках и люди с Холодной Спины. Торгрим заявил свои права на кита и запретил людям с Залива разделывать, делить и увозить тушу. Флоси потребовал от него доказательств, что Эйрик передал Энунду Деревянная Нога право на плавщину. Иначе, мол, он не отдаст кита без боя. Торгрим увидел, что сила не на их стороне, и не стал нападать. Тут с юга показалась лодка. Гребцы налегли на весла и скоро подошли к берегу. Это был Сван с Холма, что у Медвежьего Фьорда, и его работники. Приблизившись, Сван сказал, чтобы Торгрим не давал себя грабить, — а они давно уже были большими друзьями, — и предложил ему поддержку. Братья сказали, что охотно ее примут. Стали они всею силой наступать. Торгейр Бутылочная Спина первым вскочил на кита и напал на работников Флоси. Торфинн, о котором уже рассказывалось, разделывал тушу. Он стоял у головы в углублении, которое сам для себя вырезал. Торгейр сказал: — Вот тебе твоя секира!
Вот что сочинили об этой стычке: Кто о жестокой битве У Реберных Скал не слышал? Рьяно бойцы безоружные Ребрами там рубились. Асы металла лихие Швырялись ошметками мяса Метко. Да только мало Славы в делах подобных! Конечно, викингов не назовешь мирными, добродушными людьми. Однако степень озверения, до которой они дошли в этой ситуации, явно выходит за рамки нормального поведения средневековых скандинавов. Слава (то есть репутация, повышающая социальный статус в соответствии с принципом "непрямой реципрокности" (см. главу "Эволюция альтруизма" кн. 2)) была их первейшей целью, и навлекать на себя насмешки скальдов уж точно не входило в круг их жизненных задач. Саблезубые кошки помогли становлению человека?Итак, первые люди, по-видимому, были падальщиками: они питались остатками добычи крупных хищников, отгоняя от желанной мясной добычи самих хищников и других падальщиков. Увеличенный мозг и сложное коллективное поведение, вероятно, способствовали выполнению этой опасной и трудной задачи. Помимо увеличения объема мозга и коллективного поведения развивались и такие важнейшие адаптации, как двуногое хождение (вплоть до быстрого длительного бега) и орудийная деятельность. Интересную гипотезу, увязывающую воедино все перечисленные адаптации, предложил Алексей Лопатин из Московского палеонтологического института (Лопатин, 2010). Он рассмотрел развитие гоминид на фоне изменчивого животного и растительного мира — ведь для понимания движущих мотивов эволюции важно оценить, как формировалась и изменялась экологическая ниша человека. Первые гоминиды еще не были падалыциками, их диета была широка и разнообразна. Но около 2,3 млн лет назад климат стал более засушливым (см. ниже), изменились ландшафты, количество и спектр объектов питания стали другими, в новых местообитаниях появились и новые хищники. Древние гоминиды были представлены несколькими видами, так что в условиях межвидовой конкуренции приходилось вырабатывать более узкую экологическую специализацию. Два миллиона лет назад на просторах саванн паслись многочисленные стада крупных травоядных животных — зебр, антилоп. Значительный сегмент растительноядной фауны составляли гигантские свиньи и родственные им виды. Присутствовали и более крупные растительноядные животные — слоны, носороги, гиппопотамы. На них охотились хищники, как очень крупные, так и среднего размера. Наиболее эффективным охотником был мегантереон — саблезубая кошка весом около 100 кг. Более крупные и страшные хищники — лев и гомотерий (другая саблезубая кошка, весившая до 190 кг, примерно как львица) — могли охотиться даже на слонов и носорогов. Саблезубые хищники в силу устройства своего зубного аппарата оставляли после пиршества богатые остатки. Их зубы были приспособлены для раздирания толстой шкуры добычи, отрывания мышц и пожирания внутренностей, но они не могли обгладывать кости, счищать мясо с позвоночника и ребер. Так что свите падалыциков было чем поживиться. Эту свиту составляли стаи гигантских гиен и семьи древних людей. Люди конкурировали с гиенами за право первыми оказаться рядом с обильными объедками удачливого саблезубого охотника. Лопатин не первым высказал мысль о тесной экологической связи между древними гоминидами и саблезубыми кошками: эта идея достаточно популярна среди палеоантропологов (Lewis, 1997). Следуя за саблезубыми охотниками, человек должен был, во-первых, не стать обедом сам, во-вторых, успеть захватить добычу быстрее гиен. Эффективное избегание хищника и осторожность вырабатывались за счет развития мозга и коллективного поведения. Гиены предпочитают активную ночную жизнь — человек стал дневным падальщиком. Гиены не слишком быстро бегают (у них короткие задние лапы) и не очень выносливы — человеку пришлось совершенствовать технику движения и длительного бега. Благодаря этому он успевал на место оставленного пиршества раньше гиен и мог как следует поживиться.
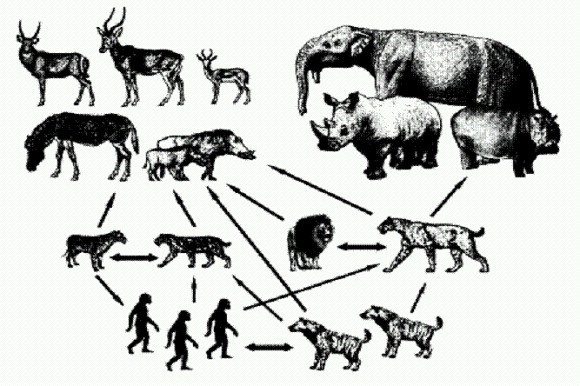
Упрощенная схема пищевой цепи млекопитающих около 2 млн лет назад. Показаны блоки среднеразмерных и крупных растительноядных животных, средних и крупных хищников и падальщиков. Черные стрелки указывают на хищнические связи, светло-серые — на следование за охотником в расчете поживиться остатками, темно-серые — на конкурентов, (из статьи А. В. Лопатина, 2010, с изм.) Судя по следам зубов на костях олдувайских копытных (2 млн лет), сначала добычей пользовался хищник, затем человек и только в последнюю очередь — гиены. Кроме того, владея орудиями разделки туш, человек мог сначала на месте разделить добычу на части, а потом унести ее в укромное место. От мастерства разделки добычи зависело выживание всей семьи. По-видимому, это и было главным стимулом развития орудийной деятельности человека.
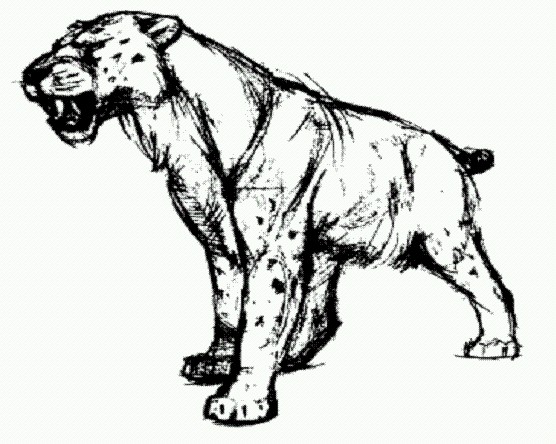
Гомотерий. Таким образом, человек занял экологическую нишу дневного, быстро бегающего, стайного падальщика. Конкурентные отношения должны были породить агрессивное поведение, учитывая размеры и свирепость гигантских гиен, которые, случалось, нападали на человека. Как можно заключить по следам зубов на костях, останки Homo erectus в пещере Чжоукоу-дянь в Северном Китае (см. ниже о заселении эректусами Евразии) погрызены и принесены туда именно гигантскими гиенами. Агрессивность в дальнейшем способствовала тому, что люди ушли от экологической ниши падальщиков и перешли к стратегии активных охотников. Для этого людям понадобились и новые навыки коллективного поведения, и усовершенствованные заостренные каменные орудия. Пока люди были спутниками мегантереонов и других крупных хищников, им достаточно было примитивных олдувайских орудий. Действительно, в Африке мегантереоны существовали до 1,5 млн лет назад — примерно до этого времени люди использовали олдувайские технологии. Затем, после вымирания этих хищников, людям пришлось менять экологическую нишу и изобретать другой орудийный набор — ашельский. В Европе мегантереоны вымерли существенно позже — 0,5 млн лет назад, тогда и древнейшие европейцы сменили набор орудий. Итак, возможно, люди и кошки были связаны гораздо раньше и намного теснее, чем обычно предполагают. Только раньше кошки "заботились" о нас, добывая нам пищу, а теперь мы заботимся о кошках. ЭректусыХабилисы не очень долго оставались "самыми человечными" из двуногих африканских приматов. В начале второго миллионолетия до н. э. к ним присоединилась другая, более продвинутая форма, отличавшаяся еще более крупным мозгом (порядка 800—1000 см3), высоким ростом и, возможно, еще более твердой и совсем уже человеческой походкой. Этих людей называют либо ранними африканскими Homo erectus ("человек прямоходящий"), либо выделяют в особый вид Homo ergaster ("человек работающий"). Как их ни называй, произошли они, очевидно, от хабилисов, причем не вытеснили их сразу, а довольно долго — примерно полмиллиона лет, до 1,5 млн лет назад — жили с ними бок о бок в Восточной и, возможно, Южной Африке. Современниками их также были Paranthropus boisei и P. robustus. Так что в начале и середине второго миллионолетия до н. э. сообщество африканских двуногих приматов оставалось не менее разношерстным, чем раньше, в третьем миллионолетии. Рост мозга и размеров тела у эректусов, вероятно, был связан с увеличением доли мясной пищи в их рационе. Впрочем, такая формулировка может навлечь на автора обвинение в ламаркизме, поэтому сформулирую точнее. Эректусы научились добывать больше мяса, то есть обеспечили себе более калорийный рацион. Скорее всего, более крупный мозг и хабилисам был бы выгоден, но хабилисы были сильнее ограничены в средствах: у них было меньше мяса на обед. Поэтому хабилисы-мутанты с более крупным мозгом страдали от голода сильнее, чем выигрывали от ума. Ведь мозг потребляет много калорий, мозговитым животным нужно больше хорошей пищи. Иными словами, мутации, увеличивающие мозг выше определенного порога, в итоге (по сумме плюсов и минусов) оказывались вредными и отсеивались отбором. С появлением более эффективных способов добычи пропитания баланс плюсов и минусов сместился (возможно, плюсы остались те же, а вот минусы уменьшились), и в результате те же самые мутации, которые для хабилисов были вредными, для эректусов стали полезными. Особи с большим мозгом стали оставлять больше потомков, и в череде поколений мозг увеличился.
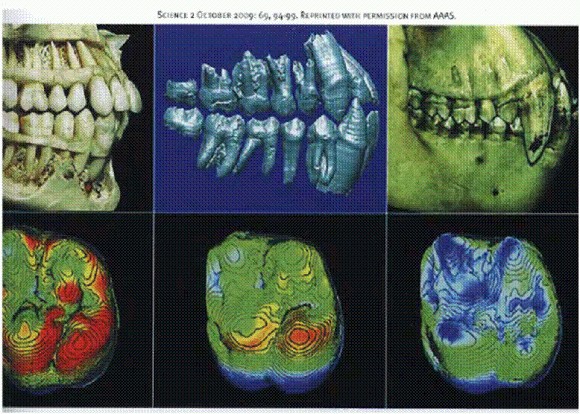
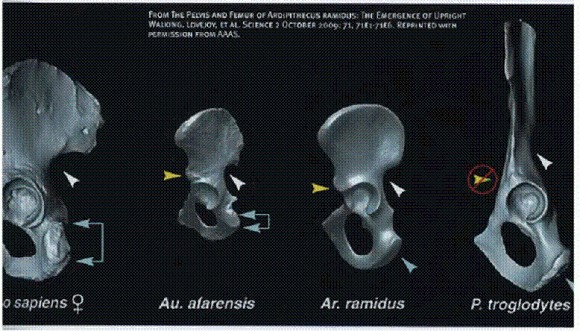
Зубы человека (слева), ардипитека (в центре) и шимпанзе (справа). Все особи — самцы. Внизу: первый моляр. Цвет отражает толщину эмали: красный — толстая эмаль (около 2 мм), синий — тонкая (около 0,5 мм). См. главу "Двуногие обезьяны".
Строение таза, слева направо: человек, Люси, Арди, шимпанзе. Таз Арди имеет промежуточное строение между шимпанзе и австралопитеком. См. главу "Двуногие обезьяны".
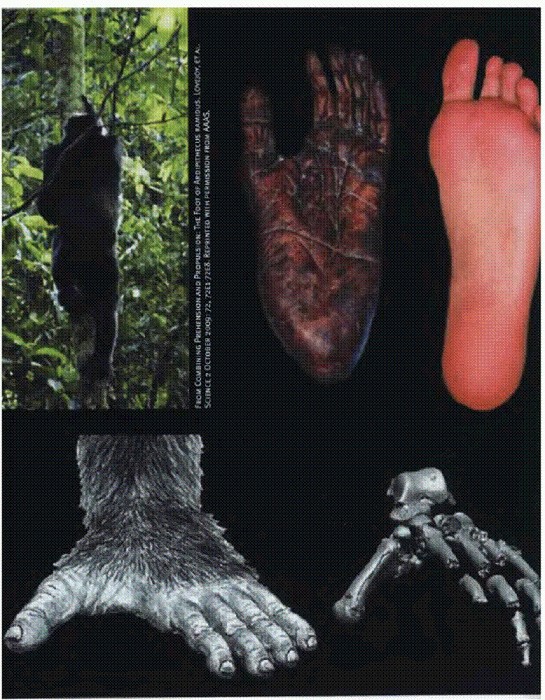
Ступни современных человекообразных обезьян хороши для лазанья и хватания за ветки, они очень гибкие и плохо приспособлены для ходьбы по земле (см. главу "Двуногие обезьяны"). Слева вверху: шимпанзе лезет на дерево. Справа вверху: ступни шимпанзе и человека. Внизу: ступня ардипитека.
Вверху: орангутаны — самец и самка с детенышем. Внизу: гориллы — самец и самка с детенышем.
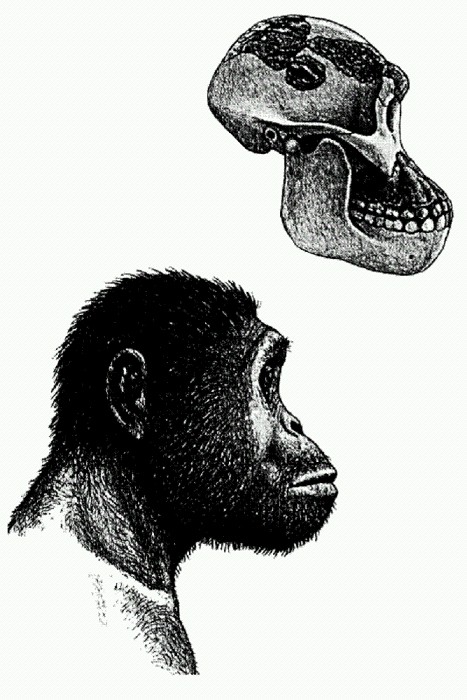
Люси, афарский австралопитек (Australopithecus afarensis). Восточная Африка, 3,2 млн лет. Реконструкция М. Л. Бутовской.
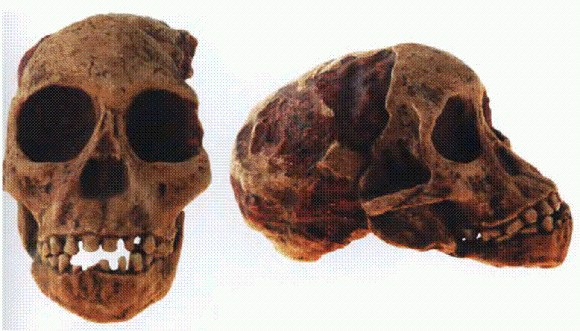
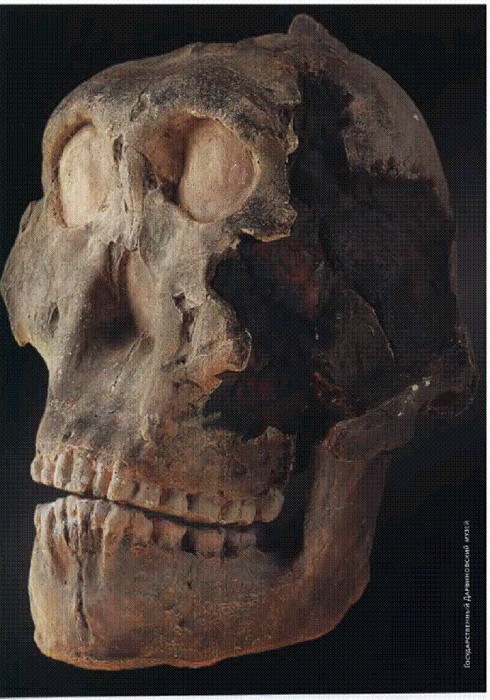
"Ребенок из Таунга": череп 6-летнего детеныша австралопитека африканского (Australopithecus africanus). Южная Африка, 2,5 млн лет. С этой находки, сделанной в 1924 году, началось изучение австралопитеков. Судя по характерным повреждениям глазниц (на фотографии они не видны), ребенок из Таунга был убит крупной хищной птицей. См. главу "Двуногие обезьяны".
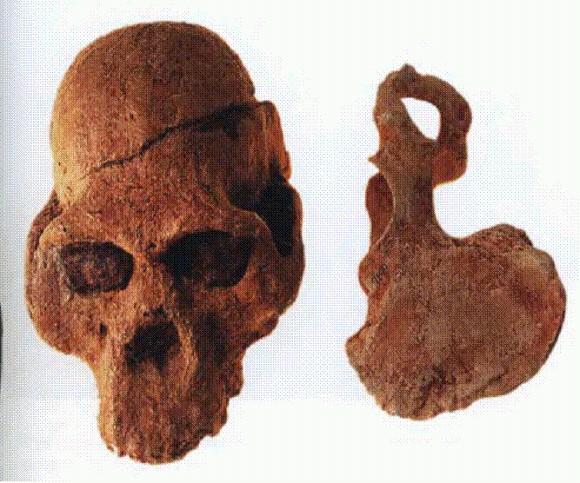
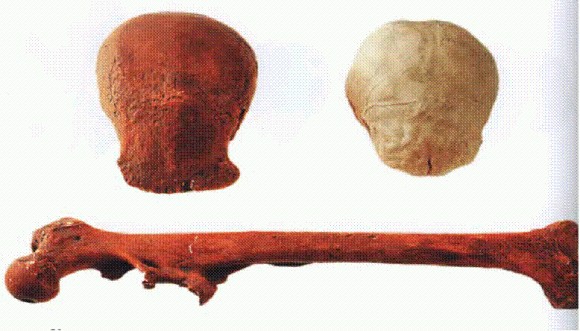
Череп и тазовая кость австралопитека африканского. Южная Африка, 3,3-2,4 млн лет.
Австралопитек африканский. Южная Африка, 3,3-2,4 млн лет. Реконструкция М. М. Герасимова.
Черепа макаки (вверху) и африканского австралопитека (Australopithecus africanus): детеныша (в центре) и взрослого (внизу). Детеныш австралопитека по некоторым признакам (выступающий лоб, укороченная лицевая часть черепа) больше похож на человека, чем взрослый представитель того же вида. См. главу "Двуногие обезьяны".
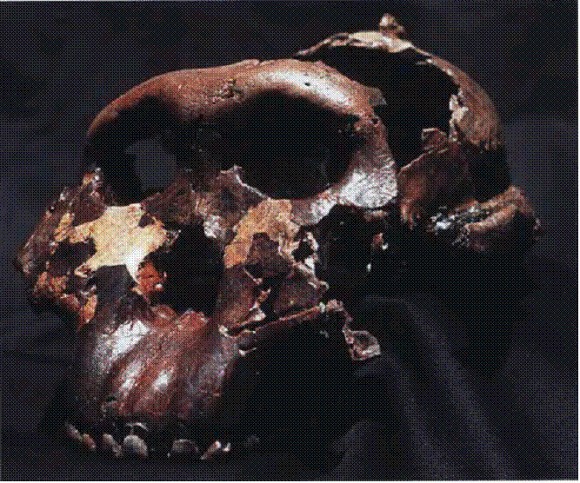
Череп "Щелкунчика" (Paranthropus boisei). В задней части черепа видны остатки костного гребня, к которому крепились мощные жевательные мышцы.
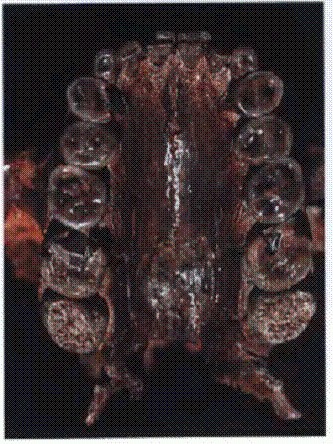
Могучие коренные зубы P. boisei. В отличие от других человекообразных обезьян, даже у самых зубастых гоминид клыки имели весьма скромные размеры. Это, скорее всего, говорит о радикальных отличиях в половом и социальном поведении, поскольку крупные клыки самцов у человекообразных играют важную роль в конкуренции за самок и служат своего рода символом мужественности. См. главу "Двуногие обезьяны".
Череп парантропа Paranthropus boisei. Восточная Африка, 2,3-1,2 млн лет.
Орудия олдувайской культуры. См. главу «Очеловечивание»
Ашельские орудия Homo erectus. См. главу "Очеловечивание".
Homo erectus: питекантроп с острова Ява, около 1 млн лет. Реконструкция М. Я. Бутовской. См. главу "Очеловечивание".
Черепная крышка, эндокран (слепок мозговой полости) и бедренная кость питекантропа с острова Ява.
Синантроп (поздний Homo erectus из Северного Китая). Около 0,5 млн лет. Реконструкция М. М. Герасимова. См. главу "Очеловечивание".
Хоббит с острова Флорес (Homo florensiensis). См. главу «Очеловечивание»
Ранний Homo sapiens, мужчина из пещеры Схул (Схул-5). Реконструкция М.М.Герасимова. См. главу "От эректусов к сапиенсам".
Древнейший костяной наконечник стрелы. Культура ховьесонс пурт, пещера Сибуду, Южная Африка, около 61 тыс. лет (см. главу "От эректусов к сапиенсам"). Похожие наконечники для стрел в историческое время изготавливали бушмены.
Продырявленные ракушки, из которых делались ожерелья. Стилбейская культура, пещера Бломбос, Южная Африка, около 71 тыс. лет. См. главу "От эректусов к сапиенсам".
Череп классического неандертальца из Ля-Шапель-о-Сен (Франция). Около 60 тыс. лет. См. главу "Другое человечество". У эректусов значительно усилился рельеф в области зоны Брока — моторного центра речи (Кочеткова, 1973). Похоже, коммуникация успешно развивалась. Увеличилась также надкраевая часть теменной доли, что может быть связано с ростом чувствительности рук и более совершенным контролем их движений (о возможной связи этой области мозга с производством ашельских каменных орудий см. в главе "Душевная механика", кн. 2). Кроме того, увеличилась затылочная доля, отвечающая за первичную обработку зрительной информации. Возможно, это было связано с дальнейшей адаптацией к жизни в открытой саванне, что, очевидно, требовало расширения кругозора.
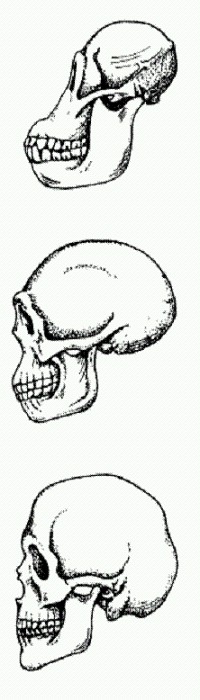
Мозг австралопитека (вверху) был вдвое меньше, чем у Homo erectus (в центре), и втрое меньше, чем у Homo sapiens (внизу). Мозг, мясо и огонь Что ни говори, а главным нашим отличием от ближайших родственников — шимпанзе или, скажем, австралопитеков — является огромный мозг. При всех очевидных преимуществах этого эволюционного приобретения оно имеет и оборотную сторону: мозг потребляет уйму калорий, которые надо где-то добывать. На содержание мозга уходит 60% всей энергии, используемой новорожденным младенцем. У взрослых расходы снижаются до 20—25%, но и это непомерно много по сравнению с другими человекообразными обезьянами (8%). При этом, однако, люди в целом потребляют не больше калорий, чем обезьяны того же размера. За счет чего достигается экономия? В случае с современным человеком ответ хорошо известен: мы питаемся пищей, которая гораздо легче усваивается, — это позволяет снизить энергетические затраты на пережевывание и переваривание. А как обстояло дело у наших предков? Антропологи давно предполагали, что наблюдаемое в эволюции гоминид поэтапное увеличение мозга должно было сопровождаться соответствующими изменениями рациона. У австралопитеков, как мы знаем, мозг был не больше, чем у шимпанзе (примерно 400—450 см3). Чуть более 2 млн лет назад он начал увеличиваться у ранних Homo (до 500—800 см3), а затем снова подрос (до 800—1000 см3) у ранних эректусов (они же эргастеры) около 1,8 млн лет назад. Последний этап быстрого увеличения мозга (до 1300—1500 см3) приходится на время между 500 и 200 тыс. лет назад, то есть на период становления двух наиболее продвинутых видов, венчающих эволюционное дерево гоминид: Homo sapiens и Homo neanderthalensis. Увеличение мозга у ранних эректусов традиционно связывают с ростом потребления мясной пищи. Действительно, есть прямые археологические свидетельства того, что мясо играло заметную роль в рационе эректусов. Например, в Кении найдены кости взрослой женщины-эректуса (экземпляр KNM-ER 1808) с патологическими разрастаниями костной ткани, характерными для гипервитаминоза А. Этот гипервитаминоз развивается при употреблении в пищу слишком большого количества животных жиров (Добровольская, 2005). Особенно легко его заработать, систематически поедая печень хищных зверей. Были ли эректусы умелыми охотниками или только собирали падаль — этот вопрос активно дискутируется, но факт остается фактом: они действительно притаскивали на свои стоянки туши животных или их части и скребли кости каменными орудиями. Но только ли в мясной пище тут дело? В 1999 году Ричард Рэнгем с коллегами выдвинули гипотезу о том, что ранние эректусы, появившиеся около 1,9 млн лет назад, уже умели готовить пищу на огне, что позволило резко снизить затраты организма на ее усвоение (Wrangham et al., 1999). Из этого предположения можно вывести множество занимательных следствий: разделение труда между полами (мужчина на охоте, женщина на кухне); рост статуса женщины-кормилицы в обществе, что можно увязать с предполагаемым уменьшением полового диморфизма у эректусов по сравнению с хабилисами; формирование ассоциативной связи между едой и сексом (Добровольская, 2005). Гипотеза Рэнгема основывалась на косвенных данных. Например, на том, что у ранних эректусов увеличился не только мозг, но и общий размер тела. Кроме того, у них уменьшились зубы. Это означает, что им теперь приходилось меньше работать челюстями. Для сравнения, шимпанзе тратят на жевание в среднем пять часов в сутки, а современные охотники-собиратели, готовящие пищу на огне, — только один час. В последующие годы Рэнгем и его коллеги провели ряд любопытных экспериментов для подтверждения этой идеи. Они кормили разных животных (например, мышей и бирманских питонов) сырым и вареным мясом. Питоны на переваривание вареного мяса тратили на 12,7% меньше энергии, а если мясо еще и пропустить через мясорубку, экономия достигала 23,4%. Энергетический выигрыш такого масштаба вполне ощутимо сказывается на конкурентоспособности животных и должен поддерживаться отбором. Группы древних людей, научившиеся готовить пищу, должны были размножаться быстрее, чем их отсталые сородичи-сыроядцы.
Мыши, питавшиеся вареным мясом, за пять недель набрали на 29% больше веса, чем такие же мыши, которых кормили сырым мясом (при одинаковой калорийности рациона). Это означает, что смена рациона у древних людей могла практически мгновенно - даже без всякой эволюции — сказаться на их размерах. Гипотеза представляется вполне логичной, но есть одна проблема: прямых свидетельств использования огня ранними эректусами пока не обнаружено. На сегодняшний день самое древнее абсолютно бесспорное кострище (с обгорелыми камнями, семенами и деревяшками) обнаружено в Палестине и имеет возраст 790 тыс. лет (см. главу "От эректусов к сапиенсам"). Правда, известно около десятка более ранних свидетельств использования огня. Например, в двух местах в Восточной Африке — в Кении (Кооби-фора) и Танзании (Олдувайское ущелье) — найдены обгорелые кремневые орудия и куски обожженной глины возрастом 1,5 млн лет, и в обоих случаях поблизости обнаружены кости Homo erectus. В Северной Кении (Чесованжа) известны следы употребления огня возрастом 1,37—1,46 млн лет. Здесь, похоже, был и очаг, обложенный камнями (Isaac, 1982). Аналогичные находки примерно того же возраста сделаны и в Южной Африке. К сожалению, во всех этих случаях исследователям не удалось окончательно доказать, что огонь, оставивший свои метки на камнях и костях, находился под контролем человека. Некоторые эксперты допускают, что это все-таки могут быть следы естественных пожаров.
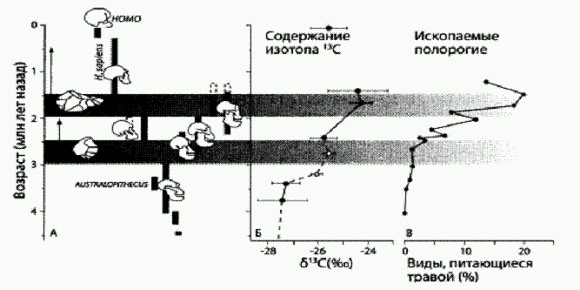
Отношение коллег-антропологов к гипотезе Рэнгема неоднозначно. Некоторые специалисты полагают, что сама по себе идея хороша, но вот с датировкой автор ошибся. Более вероятно, что люди начали готовить пищу на огне значительно позже, примерно 200—300 тыс. лет назад. Для этого времени уже есть прямые археологические свидетельства кулинарной активности наших предков. В этом случае "кулинарная революция" помогает объяснить не второй, а третий период быстрого роста мозга и уменьшения зубов — тот, который был связан со становлением сапиенсов и неандертальцев. Что же касается второго периода, связанного с появлением эректусов, то для его объяснения, вероятно, вполне достаточно увеличения доли мяса в рационе — пусть и сырого (или, может быть, слегка протухшего — этот метод "кулинарной обработки" мяса до сих пор практикуется народами Севера и, скажем, бурыми медведями). Последовательный переход ко все более легко усваиваемой пище не только снизил энергетические затраты на жевание и пищеварение, но и создал предпосылки для уменьшения объема пищеварительной системы, что тоже дало немалую экономию. У приматов обнаружена обратная корреляция между размерами мозга и пищеварительной системы. Обезьяны поумнее, как выяснилось, едят больше высококалорийной пищи (насекомые, птичьи яйца и т. п.), и пищеварительная система у них меньше. Виды с небольшим мозгом налегают в основном на легкодоступную, но малопитательную пищу (листья, плоды), и пищеварительный тракту них длиннее. Сначала предполагали, что это всеобщая закономерность, но потом оказалось, что у многих других животных данная корреляция не прослеживается. В ходе дальнейших исследований было показано, что обратная корреляция между размером мозга и длиной пищеварительного тракта выявляется только у животных с замедленным развитием, иначе говоря, с долгим детством. Долгое детство, по-видимому, является важным условием формирования крупного мозга. Что касается наших предков, то у ранних эректусов, по мнению антропологов, как раз и произошло значительное замедление роста и созревания, сопровождавшееся увеличением продолжительности жизни. Это, в свою очередь, косвенно свидетельствует о существенных изменениях образа жизни и взаимоотношений с окружающей средой, поскольку замедленное развитие и долгое детство могут позволить себе только существа, имеющие очень хорошие шансы на выживание (низкую смертность) во взрослом состоянии (Gibbons, 2007). Помимо энергетических затрат рост мозга имел еще один негативный побочный эффект: увеличение смертности при родах. Большеголовых детенышей трудно рожать. Проблема усугубляется двуногостью: антропологи утверждают, что изменения таза, которые могли бы облегчить рождение большеголовых детей, вступают в конфликт с нуждами двуногой локомоции. Если бы мозг начал расти у обезьян, передвигающихся на всех четырех, как гориллы и шимпанзе, решить эту проблему было бы легче. Но случилось так, что большой мозг "понадобился" как раз двуногим, а не четвероногим обезьянам. В качестве компенсирующей адаптации детеныши гоминид стали рождаться на более ранних стадиях развития по сравнению с другими человекообразными. Это позволило перенести большую часть роста мозга в постнатальный период. Эволюция — это бесконечный поиск компромиссов, и в ней ничего не дается даром (см. главу "Жертвы эволюции", кн. 2). Рождение недоразвитых детенышей повлекло за собой целую цепочку последствий, в том числе удлинение детства и рост нагрузки на родителей. Детеныши людей намного беспомощнее и требуют несравненно больше заботы, чем детеныши шимпанзе или горилл (Бутовская, 2004). Это в свою очередь должно было способствовать отбору на более сильную материнскую привязанность и любовь к детям. Выживаемость потомства стала еще сильнее, чем раньше, зависеть от МВП (мужского вклада в потомство). А значит, должен был усилиться отбор на способность отцов испытывать сильную эмоциональную привязанность к своему семейству. А еще — отбор на способность самок стимулировать формирование такой привязанности у партнера. Таким образом, увеличение мозга должно было способствовать эволюции любви (см. главу "Генетика души", кн. 2). Антропологи пока не пришли к единому мнению по поводу того, на каком этапе своей эволюции гоминиды начали рожать детенышей с недоразвитым мозгом. Изучение единственного прилично сохранившегося черепа младенца Homo erectus, найденного на острове Ява ("ребенок из Моджокерто"), дало противоречивые результаты. Сначала было объявлено, что мозг у юных эректусов рос скорее по "обезьяньей", чем по "человеческой" траектории. Иными словами, эректусы, подобно шимпанзе и гориллам, рождались уже с довольно большим мозгом, а период постнатального роста мозга был выражен сравнительно слабо и продолжался недолго (что затрудняло развитие речи, культуры и способности к обучению). Однако другие эксперты, переизучив череп, оспорили выводы предшественников. Возможно, эректусы все-таки развивались уже по "человеческой" траектории, то есть имели недоразвитый мозг при рождении, долгий период интенсивного постнатального роста мозга и, как следствие, долгое детство, усиленную родительскую заботу и так далее (Leigh, 2006). Проблема в том, что материала по младенцам эректусов пока явно недостаточно для окончательных выводов. Как мы уже знаем, в рационе эректусов выросла доля мясной пищи. Как им это удалось? Научились они охотиться на крупную дичь или, оставаясь падальщиками, стали успешнее конкурировать с другими падальщиками? На прошедшей в 2010 году в Париже конференции по археозоологии испанский антрополог Мануэль Домингес-Родриго доложил о результатах раскопок в местонахождении ВК (от слов Peter Bell's karongo, "овраг Питера Белла") в Олдувае. В общей сложности там было найдено более тысячи фрагментов костей животных возрастом около 1,3 млн лет, в том числе кости 30 ископаемых быков Pelorovis и двух сиватериев — ископаемых родичей жирафа. Эти животные весили не менее 400 кг. На 181 кости обнаружены царапины от каменных орудий, 172 кости раскалывались для извлечения мозга. При этом следов от зубов хищников обнаружено в общей сложности лишь 45. Самое главное, что следы от каменных орудий распределены довольно равномерно по всем частям скелета (в особенности это относится к скелетам быков). Если бы эректусам доставались лишь объедки со стола крупных хищников, а не целые туши, то царапины на костях распределялись бы иначе. Следовательно, эректусы умели охотиться на крупную дичь (Balter, 2010). Впрочем, остается еще и такой вариант: люди могли научиться отгонять хищников от убитой ими добычи и таким образом завладевать целыми тушами, даже не умея охотиться на крупных травоядных. Особенно если 1,3 млн лет назад они уже владели огнем (что, как мы помним, вполне возможно, хотя и не доказано окончательно). Не очень понятно, как эректусы могли бы завалить быка или сиватерия своими ручными рубилами (нет данных, указывающих на наличие у них копий с наконечниками или тем более луков), но ведь они могли использовать какие-нибудь западни, ловушки, деревянные колья. Хотя вопрос о времени перехода людей к активной охоте на крупную дичь остается открытым, новые находки показывают, что мясо крупных животных вошло в рацион наших предков уже 3,4 млн лет назад, а два миллиона лет спустя люди уже были в состоянии добывать целые бычьи туши. У Homo erectus была походка Homo sapiensВ 1978—79 гг. на берегу озера Туркана (в прошлом Рудольф) в Кении были найдены следы древних людей возрастом 1,43 млн лет (то есть на 2,2 млн лет моложе открытых Мэри Лики следов австралопитеков в Лаэтоли). В 2009 году там нашли еще три цепочки следов: две последовательности больших отпечатков — одна из семи, другая из двух следов и одна поменьше. Кроме того, ученые нашли и одиночные отпечатки. Цепочка из маленьких следов (два следа) находилась ниже по геологическому разрезу, этим следам примерно 1,53—1,52 млн лет. Другие две цепочки из более крупных следов немного моложе по геологическому возрасту: 1,52—1,51 млн лет. Хорошая сохранность следов позволила оценить рост, походку и особенности телосложения оставивших их людей. Походка Homo sapiens резко отличается по своей механике от четвероногого хождения других млекопитающих. Она должна способствовать амортизации толчков при шагах, увеличению подвижности тела при поворотах, ускорению бега. Амортизация толчков достигается перераспределением нагрузки на изгибы позвоночника и арки стопы; увеличение степеней свободы при движении — это результат изменения строения суставов ног. Механика шага суммирует все эти изменения, добавляя к этому увеличение эффективности движения при ходьбе и беге. Животные делают шаг, ступая на пальцы ног, затем вес тела переносится на плюсну. Человек делает все наоборот: сначала он ступает на пятку, затем максимальное давление переносит на наружный край стопы, затем на первую и пятую кости плюсны, а потом на пальцы ног. Пальцами ног человек отталкивается, делая следующий шаг. В связи с этим распределение давления при четвероногом и двуногом хождении совершенно различно. Это хорошо видно по глубине вмятин в разных частях следов. Оказалось, что походка людей, приходивших к берегу озера Туркана полтора миллиона лет назад, напоминала походку современного человека. К такому заключению ученые пришли, изучив распределение вмятин по площади следа. Размер больших следов составляет около 28 см, длина шага — от 43 до 53 см. По длине ступни можно оценить рост человека (использовались соотношения для австралийских аборигенов). Если бы такие следы оставил австралийский абориген, то он имел бы рост 1,75 ± 0,26 м. Вероятнее всего, древний пешеход принадлежал к виду Homo erectus. Следы поменьше принадлежали человеческому существу ростом около метра. Морфология следа представляет собой промежуточный вариант между австралопитеком и современным человеком. Большой палец прижат к остальным пальцам больше, чем у австралопитека, но все же меньше, чем у современного человека (угол между осью стопы и осью большого пальца у австралопитеков, эректусов с озера Туркана и современного человека соответственно 24, 14 и 8 градусов). Свод стопы уже имеется, но не настолько хорошо развит, как у современного человека. Это показывает постепенное совершенствование эффективности двуногой ходьбы и бега в линии от австралопитеков к человеку разумному. Человек, имевший такую походку и рост, как древние пешеходы с озера Туркана, мог преодолевать большие расстояния и носить тяжести. Вот так постепенно, из мельчайших деталей, и складывается реальный портрет нашего предка — человека прямоходящего (Bennett et al., 2009). Пара слов о влиянии климатаЕсли честно, я недолюбливаю "климатические" гипотезы, объясняющие эволюционные события переменами погоды. Мне было бы приятнее думать, что естественный отбор гордо и уверенно вел гоминид по пути очеловечивания без оглядки на такие пустяки, как колебания среднегодовой температуры и количество осадков. Но я вынужден пойти наперекор своим эстетическим предпочтениям, потому что факты с ними не согласуются. Конечно, изменения климата не были первопричиной появления человека. Утверждать, как это делали раньше некоторые авторы, что "появление человека было следствием наступления ледникового периода", — это уж слишком. И все же погодные условия влияли, и еще как влияли, на направленность отбора, действовавшего на древних гоминид. В других климатических условиях все могло сложиться иначе. Реальность такова, что восходящая еще к Дарвину идея о роли климатических изменений в эволюции предков человека не только подтверждается новейшими данными, но и обрастает важными новыми подробностями. Прогресс в этой области обеспечивается, с одной стороны, успехами палеоантропологии, благодаря которым эволюционная история африканских гоминид известна сегодня гораздо лучше, чем 15-20 лет назад, с другой — появлением более надежных палеоклиматических реконструкций (deMenocal, 2011). Антропологи давно предполагали, что наши предки вышли из лесов в саванну и научились ходить на двух ногах в связи с аридизацией (ростом засушливости) климата, сокращением площади лесов и распространением открытых саванных ландшафтов. Сегодня мы знаем, что уже самые древние из известных гоминид (сахелянтроп, оррорин, ардипитек), жившие от 7 до 4,4 млн лет назад, по-видимому, передвигались преимущественно на задних конечностях, хотя жили вовсе не в открытой саванне, а в негустых лесах или на смешанных ландшафтах, где лесные участки чередовались с открытыми. Переход из густых лесов на открытые пространства, так же как и совершенствование двуногой походки, происходили постепенно. Начались эти процессы, по-видимому, задолго до периода быстрой аридизации климата, распространения саванн и сокращения лесов. Этот период начался позже, 3—2,5 млн лет назад, когда гоминиды были уже вполне сформировавшейся и процветающей группой двуногих человекообразных обезьян. Об этом свидетельствуют следующие факты. ИСКОПАЕМАЯ ЛЕТОПИСЬ ПОЛОРОГИХ (BOVIDAE). К этому обширному семейству парнокопытных относятся всевозможные буйволы, быки, антилопы, газели, козлы и бараны. Полорогие составляют около трети всех находок ископаемых африканских млекопитающих (гоминиды — менее 1 %). За последние 6 млн лет в эволюции африканских полорогих было два периода наиболее быстрой смены видового состава: около 2,8 и 1,8 млн лет назад. В обоих случаях появилось много новых видов с высококоронковыми (гипсодонтными) коренными зубами, приспособленными для пережевывания жесткой саванной растительности. Доля таких видов среди африканских полорогих начала расти около 3 млн лет назад и достигла максимума 1,5 млн лет назад. По-видимому, это указывает на распространение саванных ландшафтов в этот период и на активное их освоение полорогими. Оба этапа быстрого изменения видового состава полорогих совпадают с важными событиями в эволюции гоминид. Около 2,9 млн лет назад исчезает из палеонтологической летописи вид Australopithecus afarensis (к которому относится Люси), около 2,6 млн лет назад появляются первые каменные орудия олдувайского типа, около 2,5 млн лет назад на эволюционной сцене появляются парантропы со своими необычайно мощными зубами и челюстями. Второй период смены видового состава полорогих (около 1,8 млн лет назад) примерно совпадает с появлением Homo erectus/ergaster, у которых вскоре стал быстро увеличиваться мозг. Немного позже (1,7—1,6 млн лет назад) эректусы изобрели более сложную — ашельскую технологию изготовления каменных орудий. Все эти изменения можно интерпретировать как поэтапную адаптацию гоминид к жизни в открытой саванне, сопровождавшуюся включением в рацион жесткой растительной пищи (парантропы) или мяса крупных травоядных (Homo). МУССОННЫЕ ЦИКЛЫ. Африканский климат в течение последних 5 млн лет многократно менялся с засушливого на влажный и обратно. Одной из причин этих изменений является прецессия (изменение наклона) земной оси с периодом около 25 тыс. лет. Наклон оси влияет на интенсивность муссонов, от которых зависит количество дождей, проливающихся на Африку. В дождливые периоды в Средиземном море возле дельты Нила откладываются слои сапропеля — богатого органикой ила. В последовательности из сотен таких слоев, образовавшихся за миллионы лет, прослеживается более крупномасштабная периодичность — 100- и 412-тысячелетние циклы, связанные с изменением эксцентриситета земной орбиты. Некоторые (хотя и не все) периоды более интенсивного отложения сапропеля совпадают с увеличением глубины больших восточноафриканских озер, о чем свидетельствуют глубоководные озерные отложения. Один из периодов максимальной глубины озер совпадает с появлением олдувайских орудий, а следующий такой период приходится на момент перехода к ашелю. Возможно, кратковременные дождливые периоды на фоне общего нарастания аридизации стимулировали эволюцию гоминид.
Изменения африканского климата и эволюция гоминид. По вертикальной оси — время в млн лет назад. Двумя темными горизонтальными полосами обозначены этапы "сгущения" важных событий в эволюции гоминид (А). Олдувайская обработанная галька и ашельское рубило маркируют моменты появления соответствующих каменных индустрий. Б — изменения содержания изотопа 13С в компонентах растительного воска (по результатам подводного бурения в Аденском заливе); рост этой величины свидетельствует о распространении саванной растительности. В — процент видов ископаемых полорогих, приспособленных к питанию саванной растительностью. По рисунку из deMenocal, 2011. РАСШИРЕНИЕ САВАНН. На фоне циклических колебаний влажности происходил также и направленный сдвиг в сторону аридизации климата в Восточной Африке. Его причиной было развитие периодических оледенений в высоких широтах (то самое "наступление ледникового периода"), а результатом — расширение площади саванн и сокращение лесов. Помимо эволюции полорогих, о которой говорилось выше, на усиливающуюся аридизацию указывает рост количества пыли, сдуваемой ветром с африканского континента в океан. Судя по этому показателю, аридизация началась около 2,8 и достигла максимума 1,8—1,6 млн. лет назад (deMenocal, 2004). Еще один источник данных о палеоклимате восточной Африки — соотношение стабильных изотопов углерода 13С и 12С в ископаемых растительных остатках. Саванная растительность из-за особенностей биохимических систем фотосинтеза содержит больше изотопа 13С, чем лесная. Анализ изотопного состава растительных остатков в древних почвах и в морских отложениях Аденского залива (это ближайшее к области распространения древних гоминид место, где проводилось подводное бурение) показал значительный рост содержания 13С начиная с 3—2,5 млн лет назад; максимум был достигнут около 1,5 млн лет назад. Таким образом, несколько независимых источников данных подтверждают аридизацию и распространение саванн в этот период. Все эти факты свидетельствуют о том, что климатические изменения, по-видимому, действительно оказали существенное влияние на эволюцию африканских гоминид. Значит ли это, что древние афарские австралопитеки и близкие к ним формы так и не дали бы начала ни парантропам, ни людям, если бы з млн лет назад климат в Африке не начал становиться более засушливым? Точного ответа на этот вопрос никто не знает (одним из аргументов против, на мой взгляд, являются приведенные в начале главы данные об использовании австралопитеками каменных орудий уже 3,4 млн лет назад). Но вряд ли можно сомневаться в том, что в иной климатической обстановке эволюция гоминид (а также бовид и других африканских животных) протекала бы иначе. Покорение ЕвразииРанние эректусы были первыми гоминидами, сумевшими выбраться за пределы родного африканского континента и освоить просторы Евразии. Причем произошло это уже вскоре после их появления, о чем свидетельствуют, в частности, находки грузинских археологов. Обнаружение останков "людей из Дманиси" стало одной из самых громких сенсаций в палеоантропологии конца XX века. Впервые кости древних людей, обитавших на территории Грузии 1,77 млн лет назад, были найдены в 1991 году, и с тех пор грузинские археологи, возглавляемые Давидом Лордкипанидзе, то и дело выкапывают там что-то новенькое. Сначала больше везло на черепа: их найдено и описано уже четыре. Особенно интересен один из них, принадлежащий глубокому старику (или, может быть, старухе). Обладатель этого черепа лишился почти всех зубов задолго до смерти. Находка показывает, что люди из Дманиси заботились о своих стариках. Впоследствии были опубликованы и описания посткраниального скелета. Грузинские находки интересны прежде всего тем, что это самые древние ископаемые остатки людей (и вообще гоминид), найденные за пределами Африки. В строении их черепов сочетаются примитивные признаки, характерные для древнейших представителей человеческого рода — Homo habilis (в том числе маленький мозг объемом около б00— 680 см3), и более прогрессивные черты, сближающие людей из Дманиси с типичными Homo erectus. Такая же мозаика примитивных и продвинутых черт выявилась и в строении посткраниального скелета. Древнейший достаточно полный посткраниальный скелет "настоящего" эректуса (возраст 1,55 млн лет) по всем основным признакам очень близок к нашему. Строение посткраниального скелета у хабилисов, к сожалению, до сих пор изучено слабо. Таким образом, детали преобразований скелета из того состояния, которое было у австралопитеков, в то, которое сформировалось у эректусов, проследить не удавалось, и в этом смысле грузинские находки представляют исключительный интерес. Ноги у людей из Дманиси были почти такие же длинные, как у эректусов и современных людей, и заметно длиннее, чем у австралопитеков. По-видимому, эти люди отлично бегали и могли проходить пешком большие расстояния. Об этом же свидетельствует и строение позвонков. Судя по пропорциям и форме костей, ноги у людей из Дманиси были почти человеческие, не считая отдельных примитивных черт. Руки у них, однако, были скорее как у австралопитеков, что особенно заметно в строении плечевого сустава (по этому признаку люди из Дманиси напоминают также "хоббитов" с острова Флорес, о которых речь пойдет ниже). По коэффициенту энцефализации, отражающему соотношение объема мозга и размеров тела, люди из Дманиси ближе к хабилисам, чем к эректусам. По строению позвоночника они, наоборот, ближе к эректусам. В целом по строению скелета люди из Дманиси занимают промежуточное положение между хабилисами и эректусами. Авторы находок сначала предложили выделять их в особый вид Homo georgicus, но не все антропологи с этим согласились. В принципе это дело вкуса: их можно называть примитивными эректусами, продвинутыми хабилисами или выделить в отдельный вид, а можно и вовсе оставить без названия, подчеркнув тем самым их переходный статус. Именно по такому пути и идут в последнее время многие эксперты, называя этих людей просто ранними Homo из Дманиси. Пожалуй, это лучше, чем называть их ранними эректусами: по мнению грузинских археологов, до настоящих эректусов людям из Дманиси еще довольно далеко.
"Человек из Дманиси (рассматриваесый как примитивный представитель Homo erectus в широком смысле или выделяется в особый вид Homo georgicus,). Грузия, 1,77 млн лет назад. Вопреки высказанным ранее догадкам на костях не обнаружено никаких признаков того, что их обладатели были жертвами хищников. В частности, сохранились целиком некоторые мелкие косточки, которые почти никогда не сохраняются в таком виде после трапезы хищного зверя. В Дманиси найдены не только человеческие кости, но и довольно много одновозрастных скелетных остатков всевозможного крупного и мелкого зверья. На некоторых костях сохранились царапины, оставленные каменными орудиями. Одна кость, принадлежавшая большому травоядному животному, была разгрызена крупным хищником уже после того, как люди соскребли с нее мясо. Эта находка не может служить строгим доказательством того, что люди из Дманиси уже умели охотиться на крупных животных, но она по крайней мере показывает, что они могли получать доступ к тушам раньше своих конкурентов — медведей, гиен, леопардов и саблезубых тигров (Lordkipanidze et al., 2007). Каменные орудия, найденные вместе с костями людей из Дманиси, — олдувайские, самые примитивные. Вскоре после своего появления в Грузии древние люди начали расселяться по югу Азии. На крайний юго-восток континента эректусы проникли, вероятно, около 1,5 млн лет назад. Несколько позже, около 1,2—1,3 млн лет назад, они объявились на северо-востоке Китая (судя по найденным там каменным орудиям). Не исключено, что эти две популяции — китайская и юго-восточная — были изолированы друг от друга. Обе популяции жили в районах с преобладанием степных ландшафтов и множеством крупных травоядных животных. Между ними простирались густые субтропические леса, занимавшие значительную часть южного Китая и Индокитая. Эти леса, населенные орангутанами, гиббонами, вымершими огромными обезьянами гигантопитеками и гигантскими пандами, по-видимому, были непригодны для жизни Homo erectus, предпочитавших открытые пространства.
Homo erectus. Африка, Азия, Европа, 1,9—0,4-?0,05 лет назад. Северо-китайская популяция, известная под названием синантропов, приспособилась к жизни вдали от тропической зоны (около 40° северной широты) в то время, когда периодические оледенения были сравнительно невелики и непродолжительны. И все же это ясно указывает на высокую приспособляемость эректусов по сравнению с более ранними гоминидами. Конечно, ледники никогда не доходили до Северо-Восточного Китая, но во время оледенений здесь становилось значительно суше и холоднее. В дальнейшем (около 900 тыс. лет назад) оледенения стали сильнее и продолжительнее. Северная популяция эректусов тем не менее продержалась еще несколько сот тысячелетий и закончила свое существование не ранее чем 400—350 тыс. лет назад (Shen et al., 2009). Южная популяция, приспособившаяся к жизни в экваториальной зоне, продержалась существенно дольше: последние представители Homo erectus, возможно, жили на острове Ява еще 50 тыс. лет назад (по поводу этой датировки идут споры). "Хоббиты" с острова ФлоресВ октябре 2004 года в журнале Nature было опубликовано описание неизвестного ранее вида ископаемых людей — Homo floresiensis, живших на острове Флорес (Индонезия) менее 20 тыс. лет назад (то есть совсем недавно, когда неандертальцы уже вымерли, а сапиенсы широко расселились по Старому Свету). Находка была сделана в пещере Лянг Буа. Эти странные люди, прозванные хоббитами за маленький рост (не более метра), имели крошечный мозг (около 400 см3, примерно как у шимпанзе или даже немного меньше) и при этом изготавливали довольно совершенные каменные орудия. Авторы уникальной находки указали на сходство "хоббитов" с Homo erectus. По их мнению, "хоббиты" скорее всего являются измельчавшими потомками эректусов, поселившихся на острове более 800 тыс. лет назад (судя по находкам орудий). Эта точка зрения многим экспертам с самого начала представлялась наиболее обоснованной. Однако предлагались и другие интерпретации. Некоторые авторы допускали, что Homo floresiensis произошел не от эректусов, а от более архаичных гоминид — австралопитеков, остатки которых до сих пор известны только из Африки. Другие вообще утверждали, что "хоббит" был просто-напросто необыкновенно уродливым представителем Н. sapiens, страдавшим микроцефалией в тяжелой форме. Вокруг "хоббитов" развернулась бурная дискуссия. Я расскажу о ней поподробнее, чтобы читатели могли получить общее представление о том, как палеонтропологи решают спорные вопросы при отсутствии прямых доказательств. В апреле 2006 года в Пуэрто-Рико прошло совещание Палеоантропологического общества, на котором было сделано несколько важных докладов (Culotta, 2006). Сюзан Лapсон из Университета штата Нью-Йорк сообщила о новых анатомических деталях строения скелета Н. floresiensis, подтверждающих исходную версию о происхождении этого вида от Н. erectus. Плечевая кость "хоббита", как выяснилось, существенно отличается от нашей. Если у нас головка плечевой кости повернута относительно локтевого сустава на 145—165 градусов, благодаря чему наши локти направлены назад и нам удобно работать руками перед собой в положении стоя, то у "хоббита" головка плеча повернута лишь на 110 градусов. Эта особенность сама по себе могла бы затруднить орудийную деятельность Н. floresiensis, однако ее компенсирует несколько иная, чем у нас, форма и ориентация лопатки. По-видимому, "хоббиты" могли работать руками так же эффективно, как и современные люди, однако удобное положение рук достигалось несколько иным способом, и "хоббитам" во время работы приходилось немного горбиться. А вот бросать предметы на большие расстояния они, скорее всего, не могли. Ларсон изучила также скелеты других ископаемых гоминид и обнаружила, что у единственного более или менее полного скелета Н. erectus, найденного в Кении, плечевая кость устроена так же, как у Н. floresiensis. Раньше на это не обращали внимания. Ларсон пришла к выводу, что преобразования плеча на пути от первых гоминид к современному человеку проходили в два этапа, причем "хоббит" и эректус соответствуют первому из них. В другом докладе на том же совещании Уильям Джангерс, коллега Ларсон по университету, рассказал о результатах реконструкции таза "хоббита". Хотя в ранних публикациях сообщалось о сходстве тазовых костей Н. floresiensis с таковыми австралопитеков, Джангерс обнаружил ряд продвинутых признаков, что подтверждает версию о происхождении "хоббитов" от Н. erectus. Авторы сенсационной находки считали, что найденный ими скелет принадлежал особи женского пола, однако впоследствии некоторые специалисты усомнились в этом. Джангерс сообщил, что кости конечностей других индивидуумов, обнаруженные рядом с первым скелетом, заметно меньше по размеру. Это наводит на мысль, что, возможно, у этого вида имелся половой диморфизм (мужчины были крупнее женщин). Не исключено, что скелет принадлежал мужчине, а отдельные кости конечностей — женщинам. Джангерс также сообщил, что среди найденных костей имеется и хорошо сохранившаяся ступня, которая до сих пор должным образом не исследована. Обращает на себя внимание ее крупный размер: не исключено, что "хоббиты" с острова Флорес, как и герои романа Толкина, были обладателями внушительных волосатых ступней. В целом по мере изучения "хоббитов" позиции скептиков становятся все более шаткими. Однако некоторые из них продолжают упорствовать. Например, палеоантрополог Роберт Мартин и его коллеги полагают, что мозг "хоббитов" слишком мал для того, чтобы принадлежать полноценной гоминиде таких размеров (Martin et al., 2006). "Я не уверен на 100 %, что это микроцефалия, — говорит Мартин. — Я лишь утверждаю, что мозг у него все-таки слишком маленький". Самый веский аргумент против "микроцефальной" версии состоит в том, что помимо исходного скелета в пещере Лянг Буа на острове Флорес обнаружены остатки еще нескольких индивидуумов, явно относящихся к тому же виду. По справедливому замечанию Джангерса, Мартин фактически настаивает на том, что на острове существовала целая деревня идиотов-микроцефалов. Правда, черепная коробка сохранилась только у одного экземпляра (LB1), но имеются нижние челюсти, строение которых позволяет предполагать, что у других обитателей Лянг Буа мозг был не крупнее, чем у LB1.
Homo floresiensis. Остров Флорес (Индонезия), 95—12 тыс. лет назад. Любопытно, что и после открытия неандертальцев некоторые ученые пытались доказать, что найденные кости принадлежат не особой разновидности древних людей, а людям современного типа, страдавшим олигофренией. Похоже, ситуация повторяется, причем исход дискуссии, похоже, снова будет не в пользу скептиков. Вскоре после публикации комментария Мартина и его коллег большая международная группа археологов сообщила о новом весомом аргументе в пользу того, что Homo floresiensis — не микроцефал, не пигмей и не галлюцинация, а особый карликовый вид человека. Вместе с костями девяти особей Homo floresiensis в пещере Лянг Буа первооткрыватели нашли многочисленные каменные орудия. Скептики утверждали, что эти орудия слишком совершенны, чтобы их могло изготовить существо с объемом мозга 400 см3, и что на самом деле это изделия сапиенсов — людей современного типа. Древнейшие скелетные остатки Homo sapiens, обнаруженные на Флоресе, имеют возраст 11,5 тыс. лет — они немного моложе самых молодых костей "хоббитов". Вместе с костями сапиенсов найдены куда более совершенные орудия с полированными лезвиями, а также бусины и другие артефакты, свидетельствующие о принципиально ином (верхнепалеолитическом) уровне культуры. Кроме сравнительно примитивных орудий "хоббитов" и изысканных верхнепалеолитических изделий людей современного типа на острове были найдены очень древние орудия, принадлежавшие, по-видимому, представителям вида Homo erectus, заселившим остров более 800 тыс. лет назад. Кости самих эректусов пока не найдены. Если правы скептики, считающие "хоббитов" уродливыми людьми современного типа, то орудия, найденные вместе с "хоббитами", должны сильно отличаться от продукции эректусов. Если же правы первооткрыватели "хоббитов", то Homo floresiensis — прямые потомки заселивших остров эректусов, и их орудия должны быть схожи между собой и резко отличаться от изделий Н. sapiens. Исследование, проведенное группой ученых из Австралии, Индонезии и Нидерландов, подтвердило вторую версию (Brumm et al., 2006). Впервые древние орудия (в количестве нескольких десятков) были найдены в 1994 году в 50 км к востоку от пещеры Лянг Буа. В 2004—2005 годах археологи в ходе интенсивных раскопок в этом районе добыли еще около 500 артефактов. В Лянг Буа вместе с костями хоббитов нашли 3264 каменных артефакта.
Череп "хоббита". Фото из Brown et al., 2004. Древние орудия происходят из слоя озерно-речных отложений, который в некоторых местах ограничен сверху и снизу вулканическими прослоями, что позволяет с большой точностью определить возраст артефактов. Нижний вулканический прослой имеет возраст 880 ± 70, верхний — 800 ± 70 тыс. лет. Орудия, следовательно, были изготовлены между этими датами. Каменная индустрия первопоселенцев острова была довольно примитивна — недалеко ушла от олдувайской. Большая часть артефактов сделана из низкокачественного материала — гальки вулканического происхождения, которую подбирали прямо на месте, но попадаются и изделия из более высококачественного тонкозернистого кремнистого известняка (27 артефактов) и халцедона. Материал для этих изделий, по-видимому, был принесен издалека. Технология заключалась в откалывании отщепов от ядра. Для этого просто колотили одной галькой другую или использовали в качестве молотка отработанное ядро. Отщепы получались в основном мелкого и среднего размера. Большую часть "продукции" бросали на месте изготовления и никогда не использовали. Некоторые (в основном крупные) отщепы подвергались дополнительной обработке — ретушированию. Например, край отщепа мог быть заострен путем откалывания от него более мелких отщепов. Пять орудий были обработаны особенно тщательно — от них откалывали маленькие кусочки до тех пор, пока не получили некое подобие остроконечника, который можно было использовать, например, для прокалывания дырок в шкурах. Каменная индустрия первопоселенцев острова оказалась очень похожа на изделия Н. floresiensis по основным качественным и количественным параметрам. Проколки, найденные среди древних орудий, напоминают аналогичные инструменты из Лянг Буа, а отработанные ядра из двух местонахождений во многих случаях практически неразличимы. Пожалуй, единственное существенное отличие двух наборов каменного инвентаря состоит в том, что среди находок из Лянг Буа есть 12 артефактов, раскрошившихся под воздействием огня, а среди древних орудий этого не наблюдается. Эти результаты подтверждают гипотезу о происхождении Н. floresiensis от эректусов, заселивших остров около 800 тыс. лет назад или ранее. За сотни тысяч лет спокойной жизни на острове среди экзотических карликовых слонов — стегодонов, гигантских крыс и варанов островитяне измельчали, а их технологическое развитие почти полностью остановилось. Появление на Флоресе людей современного типа — рослых верхнепалеолитических охотников с высокоразвитой каменной индустрией — обрекло маленький отсталый народец на неизбежную гибель. Впрочем, судя по легендам о маленьких лесных людях, бытующим среди современных островитян, последние "хоббиты", возможно, вымерли лишь несколько столетий назад. Дискуссия на этом не завершилась. Вскоре в журнале Proceedings of the National Academy of Sciences появилась очередная статья скептиков (Jacob et al., 2006), в которой рассматривались следующие аргументы. 1. Главный отличительный признак Н. floresiensis — маленький объем мозга (около 400 см3). Однако череп был найден только один; кости других индивидуумов не дают веских оснований утверждать, что у них тоже был маленький мозг. 2. Каменные орудия, найденные вместе с костями "хоббитов", отличаются большой сложностью. В частности, найдены мелкие лезвия, бывшие частями составных орудий. Такими технологиями владели только люди современного типа. Сходство орудий "хоббитов" с обнаруженными на острове древними изделиями эректусов, по мнению авторов, я вляется весьма поверностным. 3. Чтобы на острове сформировался новый вид людей, необходима полная изоляция. Однако нет веских оснований утверждать, что архантропы проникли на остров только единожды и в дальнейшем не было никаких контактов между обитателями Флореса и другими человеческими популяциями. Про карликовых слонов — стегодонов известно, что они проникали на Флорес как минимум дважды. А если контакты между островитянами и другими людьми сохранялись, как можно утверждать, что островитяне действительно были обособленным видом? Люди, по-видимому, могли многократно проникать на Флорес во время ледниковых максимумов, когда уровень моря падал и проливы между островами Малайского архипелага сужались или вовсе исчезали. 4. Остров Флорес слишком мал, чтобы изолированная популяция охотников-собирателей могла просуществовать на нем в течение 40 тыс. поколений. Площадь острова (14 200 км2), по имеющимся оценкам, достаточна для существования лишь 570—5700 первобытных людей, тогда как, по другим имеющимся оценкам, популяция крупных позвоночных должна начитывать как минимум 5816— 7316 взрослых особей, чтобы у нее был реальный шанс продержаться 40 тыс. поколений. 5. Карликовость сама по себе не может служить аргументом в пользу выделения "хоббитов" в особый вид, поскольку подобные вариации размеров широко распространены у многих млекопитающих, в том числе у человека. Например, африканские пигмеи вовсе не являются отдельным видом, и генетически они не изолированы от соседних племен. 6. Ни один из морфологических признаков единственного черепа "хоббита" и обеих нижних челюстей не выходит за пределы индивидуальной изменчивости, характерной для современных жителей Меланезии и Австралии. Даже те признаки, которые в первоописании были особо отмечены как "не встречающиеся у современных людей" (например, некоторые особенности строения барабанной кости), в действительности встречаются у аборигенов Австралии и недавно вымерших тасманийцев. Отсутствие у "хоббитов" подбородочного выступа также ни о чем не говорит, поскольку у пигмеев рампасаса, живущих неподалеку от пещеры Лянг Буа и специально исследованных авторами статьи, этот признак встречается весьма часто (С. В. Дробышевский по этому поводу замечает, что из опубликованных данных не понятно, как определяли отсутствие подбородочного выступа. Похоже, что это делали на глазок или даже по фотографии, а это неправильно. Для рампасасцев, как и для других меланезийцев, характерен прогнатизм (выступание вперед челюстей), поэтому сильно выступающие губы могут маскировать подбородочный выступ). 7. Главная отличительная особенность "хоббитов" — маленький мозг в сочетании с низким ростом — по мнению авторов, является следствием микроцефалии. Авторы сообщают, что микроцефалия обычно сопровождается целым комплексом других отклонений, среди которых одним из самых обычных является карликовость. Чрезвычайно высокая степень зарастания черепных швов, характерная для черепа "хоббита", могла быть первопричиной целого комплекса аномалий, включая малый объем мозга. В частности, авторы отмечают сильно выраженную асимметрию черепа, в особенности его лицевой части. Эта асимметрия выходит за пределы того, что считается клинической нормой, и говорит о том, что череп из Лянг Буа принадлежал индивиду с серьезными нарушениями развития. 8. Первооткрыватели "хоббитов" указали особенности строения зубов, которые отличают их находку от современных людей. Авторы детально разбирают эти особенности и показывают, что все они с той или иной частотой встречаются в современных человеческих популяциях, причем некоторые черты указывают на близость "хоббита" к современным флоресийским пигмеям рампасаса. 9. Рассматривая строение посткраниального скелета (то есть всех костей, не относящихся к черепу), авторы указывают на ряд черт, свидетельствующих о том, что типовой экземпляр Н. floresiensis страдал тяжелыми пороками развития. О посткраниальных костях других индивидуумов, относимых к тому же виду, авторы упоминают лишь вскользь, отмечая, что "они в целом соответствуют представлению о том, что популяция из Лянг Буа была карликовой" и, самое главное, "эти кости ничего не говорят о размерах черепной коробки их обладателей".
В большинстве учебников по антропологии можно прочесть, что подбородочный выступ - одна из самых характерных отличительных черт Homo sapiens, по которой челюсть современного человека всегда можно отличить от челюсти неандертальца или питекантропа. Однако среди пигмеев рампасаса с острова Флорес встречаются индивидуумы, лишенные подбородочного выступа. Фото из Jacob et al., 2006. Многие из перечисленных аргументов (в особенности третий, четвертый и седьмой), по-моему, весьма уязвимы для критики. Тем не менее статья стала серьезным ударом по позициям "оптимистов". Несомненно, вся аргументация скептиков мгновенно рассыплется в пух и прах, если удастся откопать на Флоресе еще один маленький череп с теми же чертами. Но этого пока не произошло. Однако в последние 2-3 года "оптимисты" явно стали одерживать верх. Это видно из того, что другие антропологи, сами не изучавшие "хоббитов", все чаще используют Homo floresiensis как материал для сравнения и основу для эволюционных гипотез. Выдвигаются и новые "оптимистические" аргументы. В 2007 году "оптимисты" опубликовали статью в Science, в которой обратили внимание на то, что до сих пор дискуссия шла в основном вокруг строения черепа и зубов, хотя кости конечностей у приматов тоже могут служить важным источником информации о родственных связях и групповой принадлежности. В частности, кости запястья Homo sapiens имеют ряд продвинутых черт, которых нет у других современных приматов. Авторы показали, что эти "человеческие" признаки характерны также и для наших ближайших ископаемых родственников: верхнепалеолитических сапиенсов (кроманьонцев) и неандертальцев, а также для Homo antecessor — вида, который считается близким к общему предку неандертальцев и сапиенсов (см. ниже). Поскольку Homo antecessor жил около 1,0— 0,8 млн лет назад, авторы полагают, что и этот комплекс признаков запястья сформировался не позже (Tocheri et al., 2007). Ученые исследовали три косточки левого запястья, принадлежащие типовому экземпляру Homo floresiensis (экземпляр LB1). Косточки были выкопаны в сентябре 2003 года, хорошо сохранились и не несут признаков патологии. Выяснилось, что все три косточки не имеют продвинутых признаков, характерных для сапиенсов и неандертальцев, и находятся в том исходном, примитивном состоянии, которое наблюдается у шимпанзе, горилл, орангутанов, австралопитеков и хабилисов. Авторы привлекли для сравнения 252 кости запястий современных людей со всех концов Земли, в том числе лиц, страдающих различными нарушениями развития скелета; 117 костей шимпанзе, 116 горилльих, 40 орангутаньих, 19 павианьих, четыре кости верхнепалеолитических сапиенсов, 13 костей неандертальцев, три кости австралопитеков и одну косточку хабилиса. Кроме того, они использовали обширные литературные данные по различным аномалиям в строении запястья, которые встречаются у современных людей. Ученые пришли к выводу, что примитивные признаки, наблюдаемые у хоббита LB1, не могут быть результатом известных болезней или аномалий развития. По их мнению, прогрессивные черты в строении запястья, характерные для современного человека, появились в промежутке между 0,8 и 1,8 млн лет назад в эволюционной линии, включающей сапиенсов, неандертальцев и их общего предка. Что же касается "хоббитов", то они произошли от каких-то более примитивных гоминид, у которых эти признаки еще находились в исходном, "обезьяньем" состоянии, как у австралопитеков и хабилисов.
Скелет кисти. Выделены косточки запястья, сохранившиеся у "хоббита" (слева направо: ладьевидная, трапециевидная, головчатая). К сожалению, кости запястья Homo erectus — вероятного предка "хоббитов" — до сих пор не найдены. Еще одна порция "оптимистических" аргументов появилась в 2009 году. Изучение эндокрана (слепка мозговой полости) единственного черепа Homo floresiensis показало, что, хотя размер мозга у "хоббита" был такой же, как у шимпанзе, его структура подверглась существенной реорганизации. В частности, изменились размеры и форма некоторых отделов коры, связанных с "высшими" мыслительными функциями. Это не обезьяний мозг, но и не мозг современного человека, страдающего микроцефалией (Ранее на сходство эндокрана "хоббита" с эндокранами австралопитеков указывал С. В. Дробышевский (2007)). Обнаруженные особенности помогают понять, как удавалось "хоббитам" при таком маленьком мозге иметь довольно высокую культуру, в частности изготавливать каменные орудия (Falk et al., 2009). Археологические данные также свидетельствуют о том, что прибывшие на Флорес около 12 тыс. лет назад современные люди, возможно, переняли некоторые приемы обработки камня у аборигенов — "хоббитов" (возраст найденных костей "хоббитов", по последним данным, составляет от 95 до 12 тыс. лет. Скорее всего, "хоббиты" дожили до прихода сапиенсов) (Moore et al., 2009). Важные факты, проливающие свет на происхождение и родственные связи "хоббитов", были получены в ходе изучения стопы LB1 (Jungers et al., 2009). По некоторым признакам стопа "хоббита" напоминает человеческую: большой палец прилегает к остальным (а не противопоставлен им, как у шимпанзе); верхняя часть свода укреплена, что позволяет эффективно отталкиваться при ходьбе; кости плюсны тоже похожи на человеческие по ряду признаков. Но в других отношениях стопа "хоббита" сильно отличается от нашей. Прежде всего, она слишком большая для человека такого роста. По относительному размеру стопы "хоббит" соответствует шимпанзе и австралопитекам, но не сапиенсам. К примитивным признакам относятся также очень длинные пальцы, кроме большого, который, напротив, укорочен (это тоже сближает "хоббитов" не с сапиенсами, а с ранними гоминидами). Стопа "хоббита" хорошо приспособлена для хождения на двух ногах, но она плохо подходит для быстрого бега. Например, ее свод не имеет особого пружинящего механизма, характерного для Homo sapiens. Длинные пальцы могли быть удобны при размеренном шаге, но были бы помехой при беге. Найденные в Кении следы Homo erectus, возраст которых составляет 1,43 млн лет (см. выше), показывают, что к тому времени наши предки уже имели вполне современную походку и строение стопы. Следовательно, эволюционные пути предков "хоббитов" и современных людей разошлись, скорее всего, раньше этой даты (если только примитивная стопа не развилась у хоббитов вторично). До сих пор наиболее вероятными предками "хоббитов" считали эректусов, которые предположительно проникли на Флорес 800-900 тыс. лет назад и постепенно измельчали, превратившись в условиях островной изоляции в особый карликовый вид людей. Новые данные свидетельствуют в пользу другой версии, которая тоже высказывалась, но представлялась менее вероятной, — о происхождении "хоббитов" от более примитивных гоминид, близких к австралопитекам или хабилисам. Эта версия, между прочим, лучше согласуется с маленьким размером мозга хоббитов. Закономерности изменения относительных размеров мозга и тела в ходе эволюции млекопитающих, в том числе приматов, хорошо изучены. На основе этих закономерностей можно заключить, что карлики метрового роста, произошедшие от сапиенсов, должны были бы иметь мозг объемом около 1100 см3, от эректусов — около 500-650 см3. Но объем мозга у LB1 — всего 400 см3, так что в этом отношении хабилисы и австралопитеки лучше подходят на роль предков "хоббитов", чем эректусы или тем более современные люди. Проблема, однако, в том, что ни австралопитеки, ни хабилисы никогда не выходили за пределы своего родного африканского континента — по крайней мере никаких фактов, указывающих на такую возможность, до сих пор не обнаружено. Чтобы добраться до далекого Флореса, примитивным африканским гоминидам пришлось бы проделать громадный путь, в том числе по таким ландшафтам, к которым они вряд ли сумели бы приспособиться. Как же предки "хоббитов" попали на Флорес? В принципе предками "хоббитов" могли быть ранние представители человеческого рода, промежуточные между хабилисами и типичными эректусами. Именно на этом этапе развития, около 1,77 млн лет назад, люди впервые вышли за пределы родного африканского континента, о чем свидетельствуют находки в Дманиси. Но объем мозга у людей из Дманиси был равен 600—650 см3 при весе около 40 кг ("хоббиты" весили примерно 30 кг). Расчеты, проведенные на основе вышеупомянутых закономерностей, показывают, что люди из Дманиси все-таки были слишком "мозговитыми", чтобы претендовать на роль предков хоббитов. Решить эту проблему неожиданным образом помогли... гиппопотамы. Правда, не современные, а ископаемые карликовые гиппопотамы, еще сравнительно недавно (тысячу лет назад) жившие на острове Мадагаскар. В условиях островной изоляции гиппопотамы, подобно предкам "хоббитов" и многим другим млекопитающим, оказавшимся в подобной ситуации, стали мельчать. При этом объем их мозга, как выяснилось, уменьшался быстрее, чем ему было "положено" в соответствии с установленными ранее закономерностями. В пределах одного вида у млекопитающих объем мозга обычно меняется пропорционально объему тела, возведенному в степень 0,25 или менее; для групп близкородственных видов этот показатель колеблется от 0,2 до 0,4. Иными словами, объем тела обычно меняется гораздо быстрее, чем объем мозга. Поэтому относительный размер мозга у карликовых форм, как правило, больше, чем у крупных. Однако в случае быстрого уменьшения размеров тела в условиях островной изоляции этот показатель, как выяснилось, может приближаться к 0,5. Это означает, что островная карликовость может вести к аномально быстрому уменьшению мозга (Weston, Lister, 2009). Если экстраполировать данную закономерность на людей из Дманиси и "хоббитов", то получается, что первые вполне могли быть предками вторых. Произошедшее при этом уменьшение мозга сопоставимо с тем, что наблюдалось у мадагаскарских гиппопотамов.
Время существования и предполагаемые родственные связи ископаемых гоминид. По вертикальной оси — время в млн лет назад. Сплошными стрелками показаны более или менее точно установленные родственные связи, пунктирными — гипотетические. Показаны два возможных варианта происхождения Homo floresiensis — от ранних эректусов и от хабилисов. По рисунку из Lieberman, 2009. Под давлением новых фактов многие эксперты, до сих пор сомневавшиеся в том, что "хоббиты" являются особым видом людей, а не выродившимся карликовым племенем сапиенсов, пересматривают свои взгляды. Среди них — Дэниел Либерман из Гарвардского университета. По мнению Либермана, наиболее вероятными предками "хоббитов" на сегодняшний день следует считать ранних эректусов, близких к людям из Дманиси, однако и хабилисов нельзя сбрасывать со счетов. Не исключено, что ранние представители человеческого рода (хабилисы, ранние эректусы и им подобные) были намного разнообразнее, а область их распространения — куда шире, чем это нам сегодня известно (Lieberman, 2009). |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Вверх |
||||
|
|
||||