|
||||
|
|
ГЛАВА 5. ЖИВОТНЫЕПредыдущая глава, посвященная эволюционным механизмам усложнения живых существ, должна была морально подготовить нас к разговору о той группе эукариот, в которой тенденция к усложнению проявилась наиболее ярко. Речь пойдет о животных. Эта группа нам ближе всех, ведь мы сами к ней относимся. Кроме того, она и по многим объективным характеристикам представляется наиболее интересной. К таким объективным показателям можно отнести и рекордное разнообразие видов (одних только насекомых описано более миллиона видов — больше, чем во всех остальных группах живых организмов, вместе взятых), и "господствующее" положение в экосистемах (в том смысле, что животные занимают самые верхние этажи трофической пирамиды), и действительно самый высокий уровень сложности среди всех живых существ. Начнем с самого начала — с того, как животные появились на свет. И снова ископаемая летописьПервые этапы эволюционного развития эукариот представлены в палеонтологической летописи весьма скудно. В течение протерозойского зона (2,5-0,54 млрд лет назад), судя по палеонтологическим данным, разнообразие и численность микроорганизмов неуклонно росли. К сожалению, далеко не всегда можно уверенно отличить ископаемых прокариот от одноклеточных эукариот. Ведь от древних микробов ничего не остается, кроме минерализованных (окаменевших) оболочек. Современные одноклеточные эукариоты обычно раз в десять крупнее прокариот, но это не абсолютное правило, и никто не знает, насколько строго оно соблюдалось в глубокой древности. Самым надежным признаком считается структура клеточной оболочки. Например, если она покрыта шипами, можно не сомневаться: перед нами представитель эукариот. Но далеко не все протисты имеют такие узнаваемые оболочки. Древнейшие ископаемые одноклеточные организмы, которых можно с полной уверенностью отнести к эукариотам, имеют возраст около 2,0-1,8 млрд лет.
Строматолиты — слоистые минеральные образования, формирующиеся в результате жизнедеятельности микробных сообществ — в течение протерозойского зона становились все более разнообразными и многочисленными. Мы можем уверенно сказать, что важнейшую роль в строматолитообразующих сообществах в этот период играли цианобактерии, среди которых встречались формы, внешне не отличимые от современных. Протерозойские строматолиты достигли высокого уровня сложности: появились формы со всевозможными ветвящимися столбиками, козырьками, разнообразной слоистостью и микроструктурой и т. п. Современные строматолиты, образуемые бактериальными матами, устроены намного проще. Из этого следует, что и протерозойские микробные маты были не в пример сложнее современных. Возможно, не последнюю роль в них играли одноклеточные эукариоты, но пока это лишь предположение.
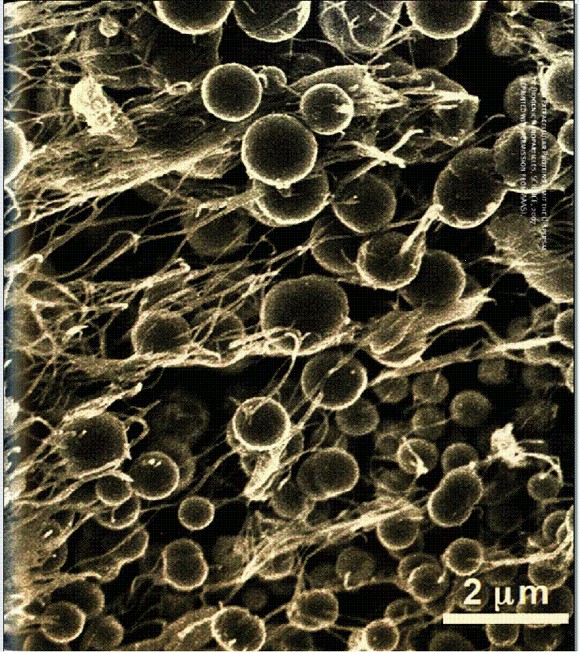
Шарики сернистого цинка в пленке сульфатредуцирующих бактерий (они на фотографии выглядят длинными нитями). Шарики получаются за счет образования связей между наночастицами сернистого цинка и цистеинсодержащими пептидами, выделяемыми клетками.
Строматолиты — слоистые образования, сформировавшиеся в результате жизнедеятельности древних микробных сообществ.
Микробиологи тоже не лишены чувства юмора. Они утверждают: "Настоящий мат должен быть трехэтажным. Если меньше трех этажей, это уже не мат, а так... биопленка". Перевернутый вверх ногами флаг африканского государства Малави помогает запомнить расположение слоев в типичном бактериальном мате. Верхний слой — зеленый из-за хлорофилла цианобактерий. Второй слой — красный или розовый из-за пигментов пурпурных бактерий. Третий слой — черный из-за сульфидов, производимых бактериями-сульфатредукторами. Красное солнце, вспыхнувшее в черном слое, можно расценивать как аллегорическое изображение появления эукариот в результате симбиоза обитателей двух нижних слоев сообщества. Правда, в современных матах слои не одной ширины: самым толстым обычно является черный слой, а самым тонким — красный.
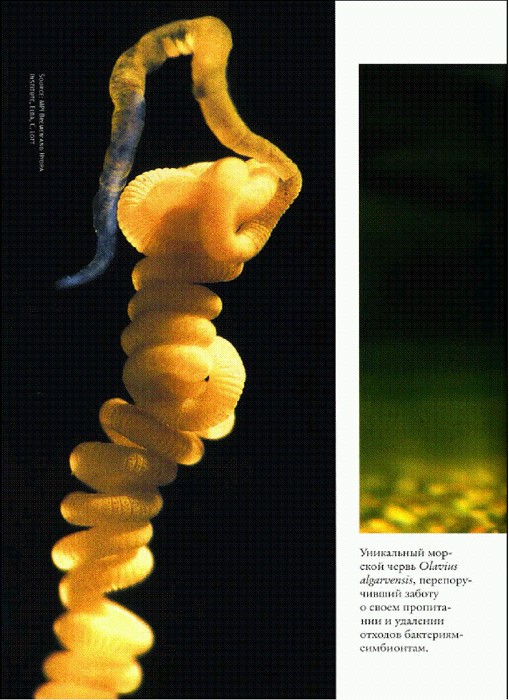
Уникальный морской червь Olavius algarvensis, перепоручивший заботу о своем пропитании и удалению отходов бактериям-симбионтам.
Аксолотль — земноводное, сохраняющее способность к восстановлению утраченных конечностей в течение всей жизни. Это сделало его излюбленным объектом биологов, изучающих механизмы регенерации.
В окрестностях озера Верхнего в Северной Америке в отложениях возрастом 1,9-1,4 млрд лет найдены спиралевидные углеродистые ленты, получившие название Grypania. Некоторые авторы считают их остатками примитивных многоклеточных эукариотических водорослей.
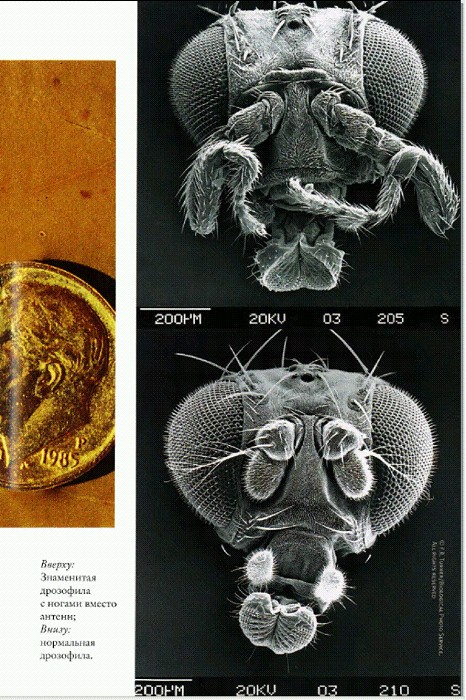
Вверху: знаменитая дрозофила с ногами вместо антенн; внизу: нормальная дрозофила.
Ход развития и строение взрослого многоклеточного животного "закодирован" в геноме примерно в той же степени и в том же смысле, в каком причудливые морозные узоры на стекле "закодированы" в структуре молекулы воды. В обоих случаях между наследственным кодом и его воплощением (генотипом и фенотипом) лежат сложнейшие, трудно поддающиеся изучению процессы самоорганизации. Эта аналогия помогает понять, почему генетики, даже имея полные тексты геномов многих видов, так медленно продвигаются к пониманию "генетических основ" сложных биологических объектов и явлений. Похоже, что первые попытки эукариот перейти к многоклеточному уровню организации начались уже в середине протерозоя. В окрестностях оз. Верхнего (Сев. Америка) в отложениях возрастом 1,9-1,4 млрд лет найдены спиралевидные углеродистые ленты, получившие название Grypania. Некоторые авторы считают их остатками примитивных многоклеточных эукариотических водорослей. Другие исследователи считают, что грипания могла быть очень высокоразвитой колонией цианобактерий. Однако есть и ряд других ископаемых находок примерно того же возраста, напоминающих многоклеточные водоросли. Хорошим кандидатом на почетное звание древнейшего животного является Horodyskia, детально изученная российским палеонтологом М. А. Федонкиным совместно с западными коллегами. Городискию нашли в отложениях возрастом около 1,44 млрд лет в Северной Америке, а также в Австралии, где возраст вмещающих пород составляет 1,4-1,07 млрд лет. По-видимому, это было колониальное многоклеточное животное, отдаленно напоминающее современных гидроидных полипов. Колония имела общий "ствол" (столон), погруженный в грунт, на котором через равные промежутки сидели однообразные округлые "особи"-зооиды. Может быть, городиския и не была настоящим животным в современном понимании этого слова. Все современные животные (Metazoa), по-видимому, представляют собой монофилетическую (имеющую единое происхождение) группу, предком которой были одноклеточные жгутиконосцы из группы хоанофлагеллят (воротничковых жгутиконосцев). Однако не исключено, что подвижные одноклеточные гетеротрофные эукариоты в течение протерозойской эры предпринимали и другие попытки перейти на многоклеточный уровень организации и превратиться таким образом в животных. Потомки этих ранних "экспериментаторов" не дожили до наших дней. Это предположение сделано по аналогии с растениями и грибами. Ведь эти два царства многоклеточных эукариот откровенно полифилетичны, т. е. каждое из них произошло не от одного, а от нескольких разных одноклеточных предков. Почему же мы должны лишать подобной вольности животное царство? На сегодняшний день считается, что одноклеточные эукариоты переходили к многоклеточности более 20 раз, однако современные животные — результат лишь одного из этих событий. Последствия всех остальных переходов к многоклеточности "достались" грибам и растениям. Возможно, городиския в какой-то мере устраняет эту несправедливость. Впрочем, некоторые авторы трактуют городискию не как животное, а как колонию одноклеточных эукариот (так называемых фораминифер). К древнейшим животным предположительно можно отнести и пармию (Parmia) — червеобразное существо, отпечатки которого обнаружены недавно М. Б. Гниловской и ее коллегами в отложениях возрастом около 1 млрд лет на северо-востоке Европейской России. Похожие отпечатки примерно такого же возраста найдены и в Китае. На некоторых отпечатках видно образование, напоминающее хоботок (такие хоботки встречаются у современных червей). Однако многие эксперты считают, что на самом деле это отпечатки водорослей, а "хоботки" — в действительности стебельки.
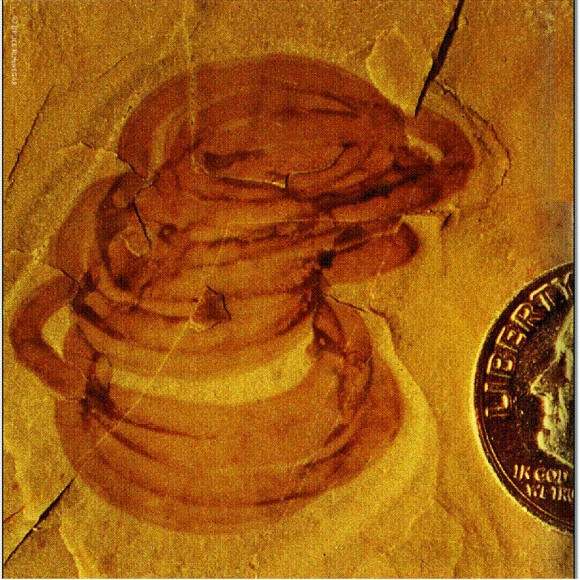
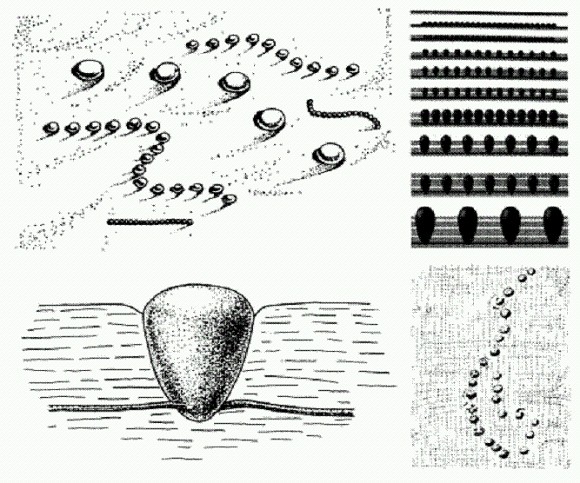
Городиския — самое древнее животное? (Общий вид, рост колонии, отдельный "зооид".) Из: М. A. Fedonkin. The origin of the Metazoa in the light of the Proterozoic fossil record // Paleontological Research. Vol. 7. № 1. E 9-41. March 31, 2003 © by the Palaeontological Society of Japan. Ископаемое Tappania возрастом 1,43 млрд лет из Австралии, по-видимому, представляет собой древнейший многоклеточный гриб, a Bangiomorpha возрастом 1,2 млрд лет из Северной Канады — практически бесспорная многоклеточная красная водоросль. Древнейшие бурые водоросли известны из отложений возрастом 1,0 млрд лет. Сравнительное изучение нуклеотидных последовательностей ДНК современных организмов показывает, что эволюционные пути растений, грибов и животных разошлись около 1,6 млрд лет назад. И это, как мы видим, неплохо согласуется с новейшими палеонтологическими данными. Однако до настоящего расцвета животных было еще далеко. В эволюции вообще было много случаев, когда между "изобретением" чего-то нового и его широким "внедрением", то есть распространением и расцветом той или иной группы организмов, проходило очень долгое время. То же самое наблюдается и в развитии человеческого общества (например, между изобретением компаса в Китае и началом его широкого применения в мореплавании прошли века). В конце протерозойского эона на Земле наступил ледниковый период, самый суровый за всю историю. Обсуждается даже "теория снежка" (snowball theory), согласно которой ледники в это время покрыли всю планету целиком. В последнее время, однако, появились данные, показывающие, что ситуация была не столь устрашающей. Тем не менее это сильнейшее похолодание, по-видимому, затормозило развитие жизни. Около 635 млн лет назад ледники наконец отступили, и начался последний период протерозоя, который называют вендским (это принятое в отечественной научной литературе название было образовано академиком Б. С. Соколовым от славянского племени вендов, или венедов, населявших южный берег Балтийского моря) или эдиакарским. Вот тут-то и началось стремительное и безудержное развитие животного мира. В глобальном масштабе происходила смена доминирующих форм жизни в океане. Микробные сообщества, в которых главными производителями органики были цианобактерии, были потеснены эукариотическими водорослями, а в портрет обитателей эпохи добавились крупные мягкотелые беспозвоночные животные.
Первопоселенцы суши. Многоклеточная жизнь зародилась в океане. На суше безраздельное господство микроорганизмов продолжалось еще примерно 150-200 млн лет после того, как в море размножились крупные многоклеточные организмы. Первыми обитателями суши, вероятно, были цианобактерии и актинобактерии. Судя по некоторым косвенным данным, они могли заселить поверхность материков еще в начале или середине протерозоя. Гетеротрофные актинобактерии образуют многоклеточные ветвящиеся структуры, похожие на грибной мицелий. Они способны объединяться с фототрофными цианобактериями в удивительные симбиотические "сверхорганизмы" — так называемые актинолишайники (см. Г. М. Зенова. Лишайники, http://evolbiol.ru/lichens.pdf. Среди первых обитателей суши были и грибы, которые тоже вступали в симбиоз с цианобактериями. Генетические и биохимические системы, развившиеся у сухопутных грибов для симбиоза с цианобактериями, позже пригодились им для "налаживания отношений" с первыми наземными растениями. Вся эта наземная микробиота постепенно готовила почву (как в переносном, так и в прямом смысле) для заселения суши растениями, которые начали осваивать пустынные берега в конце силурийского периода (около 410-420 млн лет назад). Наземные растения с самого начала жили в тесном симбиозе с почвенными грибами, без которых они, скорее всего, вовсе не смогли бы покинуть родную водную стихию (см. главу "Великий симбиоз"). Большинство вендских мягкотелых лишь весьма отдаленно напоминают современных нам животных (Об удивительной фауне вендского периода написано уже достаточно много популярных книг (см., например, свободно доступную в Интернете книгу Я. Е. Малаховской И А. Ю. Иванцова Вендские жители земли, http://evolbiol.ru/vend.htm ). О некоторых характерных вендских животных рассказано в главе "Направленность эволюции" в связи с проблемой происхождения членистоногих). Некоторые специалисты считают, что между вендской фауной и "настоящими", то есть более поздними, животными нет или очень мало прямых родственных связей. Может быть, это был еще один эволюционный эксперимент, закончившийся тупиком, а настоящие животные появились независимо от таинственных "вендобионтов"? Впрочем, по мнению ряда ведущих палеонтологов, представления о том, что вендские животные не имеют ничего общего с современным животным миром, не вполне соответствуют действительности. Загадка древних эмбрионов (Раздел написан Еленой Наймарк). До недавнего времени чуть ли не самыми загадочными и спорными из эдиакарских находок оставались многоклеточные образования из формации Доушаньтуо (Китай, возраст около 580 млн лет), похожие на ранних эмбрионов многоклеточных животных. Местонахождение Доушаньтуо найдено около 20 лет назад. С ним связаны изумительные открытия ископаемых докембрийского возраста — это самые ранние находки бесспорных многоклеточных животных: губок, кишечнополостных, билатеральных червей; известны также и отпечатки крупных водорослей замечательной сохранности. Но взрослые (хоть и микроскопические!) многоклеточные в этих отложениях редки. Самую обильную часть находок Доушаньтуо составляют похожие на эмбрионов шаровидные образования от одной клетки и более, верхний предел — около 1000 клеток. Все клетки в эмбрионе одинаковы по форме и размерам. В отличие от водорослевых клеток клетки эмбрионов не имеют толстой стенки и плотно примыкают друг к другу. Внутренней полости (бластоцеля) нет, эпителиального слоя тоже не найдено. Бластопора (эмбрионального рта) нет даже у эмбрионов из тысячи клеток. И исследователи предполагали, что эмбрионы Доушаньтуо принадлежат каким-то очень примитивным многоклеточным животным, возможно, общим предкам губок и кишечнополостных. Начало 2007 года стало переломным моментом в изучении таинственных эмбрионов. В январе американские ученые опубликовали в журнале Nature статью, в которой доказывалось, что эмбрионы Доушаньтуо — никакие не эмбрионы, а колонии гигантских бактерий. Что же навело ученых на эту мысль? Дело в том, что несколько лет назад в водах Мексиканского залива, в глубоких бескислородных слоях, были обнаружены гигантские бактерии Thiomargarita. Их размер — от 0,1 до 0,75 мм, что для бактерий поистине рекордные показатели. Эти гиганты используют для окисления сульфидов нитраты, превращая их в азот. Это так называемое "нитратное дыхание", о котором мы немного говорили в главе "Великий симбиоз" (см. раздел "Вместо выделительной системы — микробное сообщество"). Кислород нитратов переходит на сульфиды, образуя сульфаты. Запасы нитратов в клетке сконцентрированы в центральной вакуоли. Клетки Thiomargarita окружены гликопротеидным чехлом. Иногда гигантские клетки делятся, образуя скопления из двух или более соединенных клеток. На поверхности гигантских бактерий почти всегда имеются симбиотические нитчатые или сферические сульфатредуцирующие бактерии. Им для жизнедеятельности нужны как раз сульфаты, которые являются продуктом нитратного дыхания Thiomargarita. Весь этот бактериальный комплекс катализирует осаждение фосфатов — если геолог в далеком будущем нашел бы места обитания Thiomargarita, то это были бы богатейшие залежи фосфоритов. Именно таким фосфоритовым отложением и является местонахождение Доушаньтуо. Так что вполне логично предположить, что и в древних осадках фосфориты концентрировались при участии серобактерий. Также логично предположение, что этими бактериями могли быть организмы, подобные Thiomargarita. И где же они в Доушаньтуо? Единственные подходящие кандидаты — наши старые знакомые, принимаемые за яйца и эмбрионы многоклеточных. Они имеют сходные с серобактериями размеры, совершенно такое же дробление клеток в трех плоскостях, в результате которого получаются многоклеточные образования с числом клеток, кратным двум. Поверхность клеток у тех и других гладкая, те и другие окружены фосфатной оболочкой. Что касается многочисленных симбионтов на поверхности Thiomargarita, то окаменевшие нитчатые образования видны кое-где и на поверхности древних эмбрионов. Эти разительные черты морфологического сходства вместе с логичным объяснением условий накопления фосфоритового осадка заставили научное сообщество внимательно отнестись к смелой гипотезе. Однако всего через три месяца, в апреле того же года, новые ископаемые находки положили конец спорам о природе таинственных шариков. На этот раз посчастливилось китайским палеонтологам из Нанкинского института геологии и палеонтологии, работавшим в компании с Эндрю Ноллем из Гарвардского университета (США). Палеонтологи нашли множество старых знакомцев: двух-, четырех-,32-клеточных эмбрионов, но заключенных внутрь сложно устроенной оболочки.
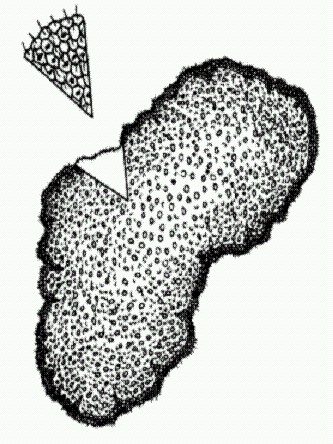
Слева — гигантские серобактерии Thiomargarita, справа — ископаемые "эмбрионы" (Nature. 2007. V. 445. Р. 198-201). Оболочка состоит из толстостенных ячеек и снаружи орнаментирована многочисленными шипами. Подобные шипастые оболочки характерны для яиц многих современных беспозвоночных. Стало ясно, что это особая эмбриональная стадия — стадия покоящихся яиц, один из способов, при помощи которого животные могут переждать неблагоприятные условия. Так что эмбрионы Доушаньтуо теперь практически безоговорочно следует признать покоящимися яйцами многоклеточных беспозвоночных. Вторая часть открытия оказалась не менее замечательной. Сами эти оболочки с шипиками, складками и бугорками на поверхности раньше находили во множестве в позднепротерозойских породах и относили к группе акритархов рода Tianzhushania. Акритархи — сборная группа проблематичных ископаемых, большинство из которых, как считалось, представляют собой оболочки цист (покоящихся стадий) одноклеточных и многоклеточных водорослей. Теперь же стало очевидно, что полые пузырьки такой формы и поверхностного рельефа — это не что иное, как оболочки яиц многоклеточных животных. Мало того, если сами эмбрионы находят в породах возрастом около 580-550 млн лет, то их оболочки под видом "акритархов" известны из самых нижних слоев формации Доушаньтуо, возраст которых — 632 млн лет. Это значит, что настоящие, бесспорные многоклеточные животные "постарели" на несколько десятков миллионов лет. И появились они в древних морях Китайской платформы всего через 3 млн лет после окончания великого позднепротерозойского оледенения. (Источники: James W. Hagadorn et al. Cellular and Subcellular Structure of Neoproterozoic Animal Embryos // Science. 2006. V. 314. P. 291-294; Jake V. Bailey et al. Evidence of giant sulphur bacteria in Neoproterozoic phosphorites // Nature. 2007. V. 445. P. 198-201; Leiming Yin et al. Doushantuo embryos preserved inside diapause egg cysts // Nature. 2007. V. 446. P. 6611-6663.) Самое примитивное животное на светеБольшинство современных типов животных впервые появляются в ископаемой летописи лишь в кембрийском периоде (это первый период палеозойской эры, начавшийся 542 млн лет назад). Однако результаты молекулярно-филогенетического анализа свидетельствуют о существенно более раннем появлении многих из них. Не стоит забывать, что палеонтология и сравнение последовательностей ДНК — не единственные источники сведений о происхождении и ранней эволюции животных. Немало полезной информации можно получить и при помощи классических методов сравнительной анатомии и эмбриологии. Вплоть до 80-х годов XX века именно эти методы служили главным инструментом для воссоздания путей эволюции животных, а палеонтология занимала почетное второе место. Сегодня на первое место вышли молекулярно-генетические методы, палеонтология осталась на втором (благодаря своему чрезвычайно бурному развитию в последние десятилетия), а анатомия с эмбриологией переместились на третье. Но своего значения они не утратили и, надеюсь, никогда не утратят. Что же говорят о происхождении животных эти классические науки? Они говорят, во-первых, что животные наверняка произошли от хоанофлагеллят — воротничковых жгутиконосцев (и это подтвердилось молекулярными данными). Во-вторых — что первые животные должны были представлять собой нечто вроде шарика (или лепешки) из двух типов клеток. Клетки наружного слоя несли жгутики и служили для движения. Внутри помещались клетки, похожие на амеб и выполняющие пищеварительную функцию. Примерно так устроены личинки низших животных — губок и кишечнополостных. Многие зоологи считали, что именно эти группы (особенно губки) представляют собой наименее изменившихся потомков общего предка всех животных, то есть являются самыми примитивными из современных представителей животного царства.
Трихоплакс — одно из самых примитивных животных — похож на медленно ползающую тонкую бесформенную пластинку. Однако на почетное звание самого примитивного животного претендовало еще одно весьма странное существо — трихоплакс. Это плоское создание, похожее на медленно ползающую кляксу, не имеет ни осей симметрии, ни мускулатуры, ни переднего и заднего концов, не говоря уже о таких сложных устройствах, как пищеварительная, нервная, кровеносная или выделительная системы. Трихоплакс по своему строению напоминает личинок кишечнополостных, и его действительно долго считали личинкой медузы. Но потом оказалось, что трихоплакс образует половые клетки и размножается половым путем. Правда, до сих пор не удалось выяснить, как идет развитие оплодотворенного яйца: в лабораторных условиях эмбрионы погибают на очень ранних стадиях. В аквариуме трихоплакс размножается бесполым путем — делится надвое или отпочковывает маленьких многоклеточных "бродяжек", у которых уже есть все типы клеток, имеющиеся у взрослого трихоплакса. Тем не менее наличие полового размножения доказывает, что трихоплакс — не чья-то личинка, а вполне взрослое, самостоятельное существо. Правда, трихоплакс мог произойти и от более сложно устроенных животных в результате неотении — редукции поздних стадий развития и перехода к размножению на стадии личинки. Это явление довольно широко распространено (один из примеров — знакомый аквариумистам аксолотль, неотеническая личинка саламандры). Окончательно решить вопрос о происхождении трихоплакса и его месте на эволюционном древе животного царства мог только тщательный молекулярно-генетический анализ. В 2006 году американские и немецкие исследователи прочли небольшую, но очень информативную часть генома трихоплакса, а именно его митохондриальную хромосому (Stephen L. Dellaporta, Anthony Xu, Sven Sagasser, Wolfgang Jakob, Maria A. Moreno, Leo W Buss, Bernd Schierwater Mitochondrial genome of Trichoplax adhaerens supports Placozoa as the basal lower metazoan phylum // PNAS. 2006. V 103. P 8751-8756.). Оказалось, что митохондриальный геном трихоплакса по своему строению занимает промежуточное положение между ближайшими родственниками животных хоанофлагеллятами и всеми остальными животными (включая губок и кишечнополостных) . Митохондриальные геномы хоанофлагеллят велики по размеру (40 и более тысяч пар нуклеотидов), содержат длинные некодирующие участки и большое число генов. Животные, напротив, имеют очень маленькие митохондриальные геномы (14-18 тысяч пар нуклеотидов) с очень небольшими некодирующими вставками и с минимальным числом генов. У трихоплакса, как выяснилось, митохондриальный геном по многим важным признакам занимает промежуточное положение. Таким образом, анализ митохондриального генома подтвердил точку зрения, согласно которой трихоплакс — самое примитивное из современных животных. Он, безусловно, не является упростившимся потомком губок или кишечнополостных, чьи митохондриальные геномы сохранили гораздо меньше примитивных черт. Простота организации трихоплакса первична. Объясняется она тем, что из всех многочисленных и разнообразных потомков "общего предка всех животных" трихоплакс изменился меньше всех. Впрочем, окончательные выводы делать пока рано. Когда эта книга уже готовилась к печати, был прочтен и основной (ядерный) геном трихоплакса. Результаты его анализа поставили под сомнение выводы, сделанные на основе изучения митохондриального генома. В ядерном геноме трихоплакса оказалось меньше примитивных черт, чем у губок. Возможно, самыми примитивными животными являются все-таки губки, хотя самые примитивные митохондрии сохранились у трихоплакса (П. Петров. Судя по полному геному, трихоплакс не так прост, как думали раньше (http://elementy.ru/news/430820).). Сравнительная геномика и ранняя эволюция животныхЕсли не учитывать редкую и плохо изученную мелочь вроде трихоплакса, то самыми примитивными животными можно назвать губок, у которых еще нет настоящих тканей, нервной системы и кишечника. Губки противопоставляются всем прочим животным — так называемым "настоящим многоклеточным животным" (Eumetazoa). Среди этих последних самыми примитивными считаются кишечнополостные, или книдарии (к ним относятся медузы, актинии, гидроидные и коралловые полипы), и гребневики. В пределах Eumetazoa книдарии и гребневики ("радиально-симметричные животные") противопоставляются двусторонне-симметричным (Bilateria). К двусторонне-симметричным относятся все прочие животные от жуков и червей до морских звезд и человека. Генетические исследования последних лет показали, что традиционные представления о примитивности книдарий, скорее всего, сильно преувеличены. В частности, оказалось, что у них, как и у высших животных, есть Нох-гены, которые в ходе индивидуального развития задают полярность зародыша и определяют план строения, в котором явно проступают черты билатеральной симметрии. Это подтвердило старую гипотезу, согласно которой общий предок Eumetazoa был двусторонне-симметричным животным. Для того чтобы составить более полное представление об этом предке, было необходимо прочесть геном представителя книдарий и сравнить его с известными геномами билатерий.
Актиния Nematostella. И вот в 2007 году американские ученые сообщили о "черновом" прочтении генома актинии Nematostella (Nicholas Н. Putnam et al. Sea Anemone Genome Reveals Ancestral Eumetazoan Gene Repertoire and Genomic Organization // Science. 2007. V. 317. P 86-94.). Геном состоит из 15 пар хромосом, имеет размер около 450 млн пар нуклеотидов (в 100 раз больше, чем у кишечной палочки, и в 6 раз меньше, чем у человека) и содержит примерно 18 000 белок-кодирующих генов, что вполне сопоставимо с другими животными. Для каждого гена актинии исследователи пытались найти аналоги в геномах билатерий: человека, дрозофилы, круглого червя, рыбы и лягушки. Если аналог (то есть похожий ген) находился, исследователи делали вывод, что соответствующий ген имелся у общего предка Eumetazoa. Таким образом удалось составить довольно полное представление о генном репертуаре этого загадочного предка. Оказалось, что репертуар этот был весьма широк и включал не менее 7766 генных семейств, сохранившихся и у книдарий, и у билатерий. Человек унаследовал не менее 2/3 своих генов от общего с актинией предка; сама актиния — примерно столько же. Муха и круглый червь унаследовали от общего предка с актинией лишь 50% и 40% генов соответственно. Выяснилось, что в эволюционных линиях позвоночных и книдарий было потеряно меньше исходных генов и меньше приобретено новых, чем в линии, ведущей к круглым червям и насекомым. Однако нужно иметь в виду, что применяемые методики не позволяли отличить действительную потерю гена от его изменения "до неузнаваемости". Поэтому в целом можно лишь заключить, что в линии позвоночных геном изменился меньше, чем в линии первичноротых, куда относятся муха и червь. Из этого следует неожиданный вывод: геном человека оказался в целом гораздо более похожим на геном актинии, чем геномы мухи и червя. Сходство затрагивает не только набор генов, но и порядок их расположения в хромосомах. Около 80% генов общего предка Eumetazoa имеют явные аналоги за пределами животного царства. Это значит, что они были унаследованы животными от одноклеточных предков (как мы помним, ими были воротничковые жгутиконосцы — хоанофлагелляты). Получается, что геном на удивление мало изменился при становлении животного царства. Среди оставшихся 20% генов, аналогов которых нет у одноклеточных, имеется большое число ключевых регуляторов развития. Примерно четверть этих новых генов (то есть 5% от общего числа) содержат участки или функциональные блоки, встречающиеся у одноклеточных, но в других комбинациях. Это указывает на один из основных путей создания новых генов: они формируются из старых путем перекомбинирования фрагментов. Как и следовало ожидать, значительная часть "новых" генов Eumetazoa выполняет функции, непосредственно связанные с теми новшествами, которые появились у животных на организменном уровне. Это прежде всего гены, отвечающие за межклеточные взаимодействия и передачу сигналов, за перемещения клеток, регуляцию их деления и другие процессы, играющие ключевую роль в ходе индивидуального развития животных. Нох-гены — регуляторы развития животных (Раздел написан Еленой Наймарк). Ключевую роль в эволюции животных играли изменения сравнительно небольшого числа генов — регуляторов индивидуального развития, от которых зависит план строения организма. Главными из этих генов-регуляторов являются так называемые Hох-гены. Прошло уже больше 50 лет с тех пор, как американский генетик Эдвард Льюис с удивлением рассматривал мутантную плодовую мушку, у которой на голове вместо антенн выросли ноги. Эта странная мутация получилась, когда ген, ответственный за формирование грудных конечностей, включился не в то время и не в том месте. А Эдвард Льюис (вместе с Кристианой Нюссляйн-Волхард и Эриком Вишаусом) получил в 1995 году за исследование этих механизмов эмбриогенеза Нобелевскую премию по физиологии и медицине. Так было открыто семейство Нох-генов, отвечающих за правильное формирование частей тела у животных. Работа этих генов казалась чудом: вот оно, решение великой загадки, как из сборища одинаковых эмбриональных клеток в правильном порядке дифференцируются ткани и органы и в результате получается сложный организм. Необходимо только в нужный момент включать правильный Нох-ген. Регуляторные Нох-гены у дрозофилы располагаются в хромосоме в довольно строгом порядке, приблизительно в том самом, в котором происходит дифференцировка основных частей тела двусторонне-симметричного (билатерального) животного. Сначала у раннего эмбриона начинают работать гены, отвечающие за строение органов на голове, затем на груди, затем гены начинают оформлять и хвостовую часть. Похожие гены были найдены у мыши и у человека. Даже у этих высокоорганизованных существ они выполняют ту же работу: отвечают за порядок эмбрионального развития. Открытие сходных Нох-генов у разных типов животных заставило зоологов и эмбриологов по-новому взглянуть на индивидуальное развитие животных и его преобразования в ходе эволюции. Стало ясно, что, изменив всего лишь один ген или время его включения, можно трансформировать, дублировать, удалить или перенести в другое место сразу целый орган, сохранив при этом общий план строения. Кроме того, ученые получили новый мощный инструмент для эволюционных построений — семейство гомологичных (происходящих один от другого) генов, присутствующих у всех животных. Все гипотезы о происхождении и ранней эволюции животных теперь включают и этот пласт информации. Нох-гены располагаются на одной или нескольких (до четырех) хромосомах, обычно тесными группами (кластерами), внутри которых сохраняется более или менее строгий порядок: "головные" гены впереди, "хвостовые" — сзади. У более примитивных представителей многоклеточных, таких как гребневики (Ctenophora) и кишечнополостные (Cnidaria), этих эмбриональных регуляторных генов только четыре, у млекопитающих их уже 48. Семейство Нох-генов подразделяется на 14 классов. Считается, что эти 14 классов возникали путем дупликации (удвоения) одного или немногих исходных генов. "Дубликаты" затем мутировали и обретали новые функции. У примитивных кишечнополостных и гребневиков имеется всего 4 класса Нох-генов, у предполагаемого общего предка двусторонне-симметричных животных их должно было быть по крайней мере 8, у млекопитающих присутствуют все 14 классов. Принцип работы всех Нох-генов одинаков. Все они являются транскрипционными факторами (регуляторами транскрипции — "прочтения" генов), то есть их функция состоит во "включении" или "выключении" других генов. В результате работы Нох-факторов запускается каскад реакций, приводящий к появлению в клетке нужных белков.
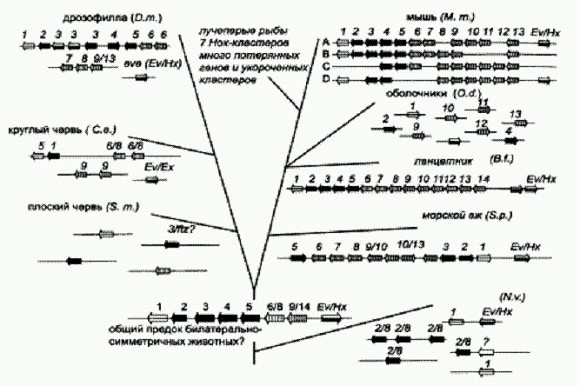
Обобщенная схема эволюции основных групп многоклеточных животных. Внизу показано предполагаемое строение Нох-кластера у общего предка билатеральных животных. У него предположительно имелось 8 Нох-генов в едином кластере. Гены нарисованы разными цветами и пронумерованы. Цифры дробью указывают, что данные гены совмещены, то есть еще не разошлись в ходе эволюции. Некоторые гены показаны одинаковым цветом, это означает большое сходство в их нуклеотидных последовательностях. Стрелочки указывают направление считывания генов на ДНК. В последние годы представления о Нох-генах существенно изменились и расширились. Этому способствовала расшифровка ДНК-последовательностей Нох-генов у многих групп животных: кольчатых, плоских и круглых червей, иглокожих, членистоногих, оболочников, ланцетников, не говоря уже о млекопитающих. Под тяжестью новых данных обрушилось представление об упорядоченном расположении Нох-генов в хромосомах. Выяснилось, что, например, у иглокожих первые три Нох-гена располагаются прямо перед последним (14-м), а начинается кластер с пятого гена. У круглых червей и оболочников Hох-гены вообще не образуют кластеров и их порядок в хромосомах не соблюдается вовсе. Это говорит о том, что правильный порядок экспрессии ("включения") Нох-генов в различных частях эмбриона не обязательно должен соответствовать порядку расположения этих генов в хромосоме. Последовательность включения Нох-генов зависит помимо "места под солнцем" от ряда дополнительных факторов, которые сейчас активно изучаются (но выяснить пока удалось немногое). Число классов Нох-генов в разных типах животных оказалось разным. Завораживающее сходство Hох-кластеров у мухи и человека тускнеет, лишь только к ним прибавляется еще какая-нибудь группа двусторонне-симметричных животных. Так, у круглых червей (нематод) найдено всего 5 классов из 14, у оболочников — 9, зато у ланцетника — все 14. У паразитических трематод (класс плоских червей) обнаружено всего 4 Нох-гена на двух хромосомах, и между этими четырьмя разбросаны другие гены, не связанные с Hох-семейством. И вот вопрос: почему у двусторонне-симметричных плоских червей Нох-генов столько же, сколько и у радиально-симметричных кишечнополостных и гребневиков? У кишечнополостных — вроде бы понятно: недостающих четырех классов генов еще не было, они образовались только после того, как появился гипотетический предок двусторонне-симметричных животных. Но у червей-то куда делась половина предковых регуляторных генов, зачем они избавились от них? Или, может быть, этой недостающей половины не было и у предка? Ответа пока нет. Ясно только, что даже при потере большого числа Нох-генов оставшиеся могут организовать эмбриональное развитие сложного билатерального существа. Зато стали известны детали регуляции самих регуляторных Нох-генов. Между Нох-генами расположены участки ДНК, прежде считавшиеся бессмысленными. В действительности, как оказалось, с них считываются короткие молекулы регуляторных РНК. Некоторые из них усиливают или ослабляют экспрессию (работу) самих Нох-генов, некоторые косвенно влияют на работу других транскрипционных факторов. В экспериментах показано, что эти микроРНК могут регулировать как соседний, так и отдаленный Hох-ген. Так что семейство Нох-генов, главное из главных среди генов-регуляторов, само не имеет полной власти в своем хозяйстве. За ним следят мелкие "выскочки" РНК, способные видоизменить экспрессию гена и тем самым замедлить или ускорить формирование органа. (Источник: Derek Lemons, William McGinnis. Genomic Evolution of Hox Gene Clusters // Science. 2006. V. 313. P. 1912-1922.) Пожалуй, самый главный вывод, который можно сделать на основе анализа генома актинии, состоит в том, что уже самые первые представители животного царства обладали весьма сложным и совершенным "набором рабочих инструментов", то есть генов, который позволил создать огромное разнообразие сложных многоклеточных организмов, внося лишь небольшие изменения в базовую программу развития. Более того, эти изменения, судя по всему, вносились не в "архитектуру" программы, а только в ее "настройки". Базовая генетическая программа поведения клетки, имевшаяся уже у первых животных, оказалась столь удачной и гибкой, что дальнейшая эволюция животного царства — в том числе и прогрессивная эволюция, связанная с усложнением организма, — уже почти не требовала радикальных изменений самой программы. Достаточно было лишь небольших изменений в "настройках". Менялись, в частности, регуляторные участки ДНК, от которых зависит тонкая настройка работы генов-регуляторов. "Главный секрет" животных. На интуитивном уровне мы привыкли относиться к царству животных как к чему-то огромному и чуть ли не бесконечно разнообразному. Но в последнее время все больше появляется фактов, которые показывают, что в действительности животные (Metazoa) представляют собой весьма специфическую, компактную и генетически однородную группу организмов. Прочтенный геном актинии — яркое тому свидетельство. По-видимому, "самый большой секрет" животных, та генеральная идея, на которой основаны их строение и эволюция, заключается в особой технологии построения сложного организма из множества изначально одинаковых модулей — клеток. Суть технологии в том, что благодаря деятельности ряда ключевых генов — регуляторов развития (в том числе Нох-генов) между делящимися клетками складывается сложная система взаимоотношений, клетки обмениваются сигналами, градиенты концентраций регуляторных веществ задают симметрию и план строения развивающегося организма, и все эти факторы вместе направляют процесс самоорганизации, самосборки сложного многоклеточного существа из генетически идентичных (то есть изначально одинаково "запрограммированных") клеток. Очень важно помнить, что геном работает не на уровне организма, а на уровне клетки. По сути дела он реально кодирует лишь биохимию и поведение одной клетки. Никакой "программы развития организма" в оплодотворенном яйце нет: там есть программа поведения клетки, не более. Что же касается "программы развития", то она самозарождается из взаимодействия делящихся клеток уже в ходе самого развития (примерно так же, как это происходит у Protozoon). Развитие червей начинается с хвоста. Нематода (круглый червь) Caenorhabditis elegans — замечательный модельный объект для изучения индивидуального развития животных, равно как и для многих других исследований. Достаточно сказать, что в 2002 году Сиднею Бреннеру, который первым начал использовать этого крошечного червя в качестве модельного объекта в генетике и эмбриологии, была присуждена Нобелевская премия "за открытия в области генетического регулирования развития человеческих органов". Работа, конечно, проводилась на черве, но выводы оказались справедливыми и для человека — вот вам и еще одно доказательство генетического единства животного царства. Генетики и эмбриологи выявили целый ряд генов, влияющих на развитие червя, причем многие из этих генов имеются также и у других животных, включая человека. Как мы уже знаем, система генетической регуляции развития, включающая Hох-гены и ряд других генов-регуляторов, в общих чертах сходна у всех животных. Однако ученые пока еще очень далеки от полного понимания тех удивительных механизмов, которые заставляют дробящееся яйцо превращаться не в комок клеток, а в сложно и тонко организованное многоклеточное существо.
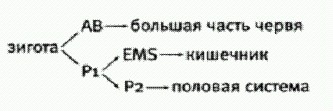
Червь Caenorhabditis elegans — любимец генетиков и эмбриологов. Одно из главных достоинств С. elegans — прозрачность. Под микроскопом хорошо видны на просвет все внутренние органы и даже отдельные клетки. Чтобы увидеть во всех подробностях, как из оплодотворенной яйцеклетки развивается червячок с пищеварительной и нервной системами, мускулатурой, органами размножения, достаточно просто положить яйцо С. elegans на предметный столик микроскопа и наблюдать за ним в течение 14 часов — именно столько продолжается эмбриональное развитие этого животного. Генетический анализ позволяет лишь выявить гены, необходимые (но вовсе не обязательно достаточные) для тех или иных аспектов дифференцировки клеток. Например, мутации Нох-генов могут приводить к причудливому перемешиванию признаков, характерных для разных отделов тела (на голове дрозофилы могут вырасти ноги, на заднем сегменте груди — дополнительная пара крыльев). Известны гены, мутации в которых приводят к полному нарушению процессов клеточной дифференцировки, так что вместо нормального эмбриона действительно образуется бесструктурный комок клеток. Но чтобы понять, как все эти гены вместе руководят развитием, необходимы эмбриологические эксперименты. С. elegans — идеальный объект для таких исследований. Развитие этого червя прослежено во всех деталях — от оплодотворенного яйца до взрослого организма. В точности известна судьба каждой зародышевой клетки (бластомера); известно, из какого бластомера образуется каждая из клеток взрослого червя. Надо сказать, что развитие круглых червей, в отличие от многих других животных, очень строго определено с самого начала. Даже на стадии двухклеточного эмбриона его бластомеры отнюдь не одинаковы. Разделив их, мы не получим близнецов, как это бывает у позвоночных. Более крупный передний бластомер АВ дает большую часть тела, мускулатуру, нервную систему; задний бластомер P1 — меньшую часть тела, включая половые органы и кишечник. Таким образом, судьба этих бластомеров известна уже на двухклеточной стадии. Она предопределена изначальной полярностью яйцеклетки: одному бластомеру достается больше каких-то регуляторных (сигнальных) веществ, другому меньше. При этом, однако, бластомеры вовсе не являются замкнутыми системами, изначально запрограммированными на какой-то определенный путь развития. Они не могут правильно развиваться без контакта с другими бластомерами; они обмениваются между собой разнообразными химическими сигналами, корректируя свое поведение в соответствии с переменами в клеточном окружении. Маркус Бишофф и Ральф Шнабель из Института генетики Технического университета в Брауншвейге задались целью выяснить, какой механизм отвечает за придание эмбриону передне-задней полярности. В экспериментах им чрезвычайно помогло то обстоятельство, что с ранними эмбрионами С. elegans можно обращаться как с миниатюрным живым конструктором. Бластомеры можно разделять и перекомбинировать произвольным образом и даже составлять химерные эмбрионы из клеток разных особей. До взрослого червя такие конструкции, как правило, дорасти не могут, но все-таки умирают не сразу и некоторое время развиваются. Исследователи отделили у четырехклеточных эмбрионов две передние клетки ("дочки" бластомера АВ), из которых в норме должен развиться почти весь червяк. Лишенные контакта с задними бластомерами ("дочками" бластомера P1, которые называются EMS и Р2), эти клетки делились хаотически и превращались в аморфный комок без всяких признаков передне-задней полярности.
Естественно было предположить, что задние бластомеры выделяют какое-то сигнальное вещество, упорядочивающее деление передних клеток. Так оно и оказалось. "Поляризующим центром" зародыша, как показали эксперименты, служит бластомер P2 (из него впоследствии развивается половая система). Достаточно приложить клетку P2 к потомкам АВ хотя бы на пять минут, чтобы клетки "поляризовались". После этого потомки АВ делятся не случайным образом, а преимущественно в определенной плоскости, так что в результате из них получается не комок, а удлиненный червеобразный зародыш с выраженным передним и задним концом. Задним концом всегда становится та точка, к которой прикоснулась клетка Р2.
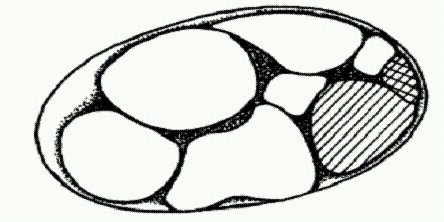
Зародыш С. elegans на стадии 8 клеток. Очевидно, сигнальное вещество, выделяемое клеткой P2, влияет на ориентацию плоскости деления у дробящихся бластомеров. Сила этого вещества такова, что, прикладывая к клеткам два Р2-бластомера в разных местах, ученые получали причудливых монстров с двумя хвостами или L-образно изогнутые зародыши с двумя взаимно перпендикулярными передне-задними осями. Каждая точка контакта с Р2 превращалась в хвост! Теперь надо было выяснить, что это за вещество. Ранее было установлено, что на ориентацию плоскости деления клеток оказывает влияние сигнальный белок Wnt, один из важнейших многофункциональных регуляторов развития у животных. Этот белок в иерархии регуляторов стоит в некотором смысле даже выше Нох-генов. Одни клетки выделяют белок Wnt, другие воспринимают его при помощи специальных белков-рецепторов, и это приводит к активизации других генов-регуляторов, в том числе Нох-генов. Чтобы проверить, действительно ли бластомер Р2 поляризует остальные бластомеры при помощи белка Wnt, исследователи использовали мутантные Р2-бластомеры с выключенным геном Wnt. Оказалось, что такие Р2-бластомеры поляризующим действием не обладают. Таким образом, природа сигнала была установлена. Оставалось понять, каким образом сигнал передается от одних клеток к другим. Исследователи обнаружили, что клетки, "поляризованные" Р2-бластомером, сами приобретают способность поляризовать другие клетки. Однако если у них выключен ген Wnt, этого не происходит. Из этого авторы сделали вывод, что передача сигнала происходит не путем диффузии сигнального вещества, производимого Р2-бластомером, из клетки в клетку (как у модельного существа Protozoon из прошлой главы), а по принципу эстафеты. Поляризованные клетки сами начинают производить белок Wnt и таким образом поляризуют своих соседей. Дополнительные эксперименты показали, что оставшаяся часть зародыша (та, что получается из бластомера EMS) тоже поляризуется благодаря контакту с Р2. Таким образом, бластомер Р2 оказался главным организующим центром развивающегося зародыша, определяющим его передне-заднюю полярность. Это важное открытие германских эмбриологов, а также разработанные ими методики сложных манипуляций с бластомерами открывают новые захватывающие перспективы перед учеными, стремящимися проникнуть в тайну развития животных. Данная работа очень наглядно показывает, каким образом довольно простые системы химического "общения" между клетками зародыша позволяют им "самоорганизоваться" в сложный многоклеточный организм. (Источник: Marcus Bischoff, Ralf Schnabel. A Posterior Centre Establishes and Maintains Polarity of the Caenorhabditis elegans Embryo by a Wnt-Dependent Relay Mechanism // PLoS Biology. 2006.4(12): е395.) Нужны ли эмбрионам гены?Имеется еще одно странное обстоятельство, затрудняющее понимание природы индивидуального развития животных. Оно состоит в том, что ранние этапы онтогенеза, как правило, идут вообще без участия генов, при полностью отключенном геноме. Задумаемся, что это значит. Ведь принято считать, что индивидуальное развитие — это постепенная "реализация" той генетической информации, которая заключена в геноме оплодотворенной яйцеклетки (зиготы) и которая в итоге окончательно "воплощается" в строении взрослого организма. Все понимают, что путь от генотипа к фенотипу труден и извилист, но лишь немногие эволюционисты всерьез пытаются объяснить ключевые закономерности эволюции особенностями тех сложнейших процессов самоорганизации, которые составляют суть онтогенеза (уж слишком трудна задача). Поэтому обычно проблему пытаются упростить, сведя все к вопросу о том, каким образом те или иные изменения генотипа (например, мутации) могут отразиться на процессе развития зародыша. Геном традиционно рассматривается как активное начало (в нем все "закодировано", он "руководит" развитием). Развивающийся зародыш, напротив, считается чем-то вроде пассивного "результата" деятельности генома. Дело несколько осложняется тем, что сам геном в процессе онтогенеза явно не самодостаточен: в разных клетках эмбриона одни гены включаются, другие выключаются в строгой последовательности, определяемой, в частности, химическими сигналами, которыми обмениваются между собой клетки и ткани зародыша. Кто кем управляет, становится не совсем ясно. Некоторые теоретики по этому поводу даже заявляют, что геном — это не "программа развития зародыша", а скорее некий набор инструментов, которыми зародыш пользуется (или не пользуется) по своему усмотрению.
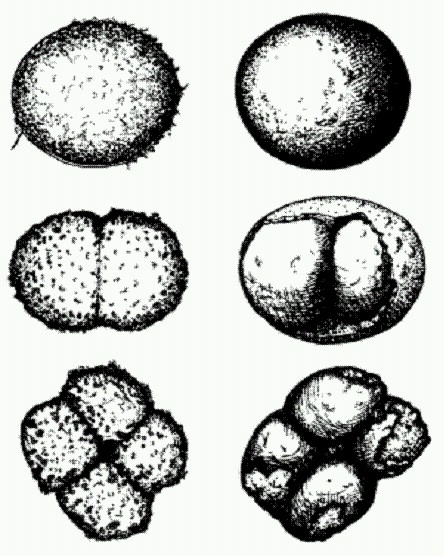
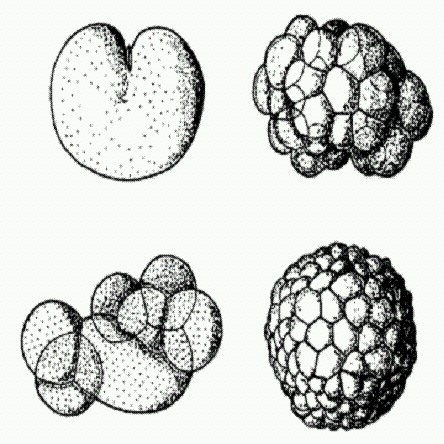
Делящаяся яйцеклетка (слева наверху), морула в виде комка клеток (слева внизу), бластула в виде полого шарика (справа внизу и вверху). На всех этих стадиях развития животных геном обычно не функционирует. На рисунке — развитие медузы Aequorea. Для управления собственными генами зародыш использует множество разных механизмов: это и регуляция транскрипции (считывания генов) при помощи малых РНК и специальных регуляторных белков — транскрипционных факторов {Нох-гены, как мы помним, тоже кодируют транскрипционные факторы), и особые "эпигенетические" механизмы, о которых мы подробнее поговорим в главе 8. Еще одно обстоятельство, которое делает сомнительным тезис о полной и однозначной обусловленности онтогенеза геномом зиготы, состоит в том, что, как уже было сказано выше, у подавляющего большинства животных на ранних стадиях развития геном вообще не функционирует. Он просто-напросто отключен, все гены молчат и матричные РНК ("считываемые" с генов матрицы для синтеза белка) не производятся.
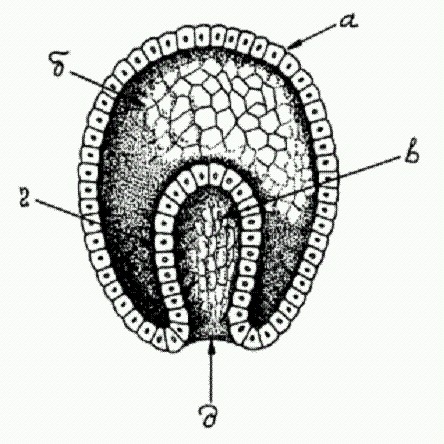
Типичная гаструла многоклеточного животного, а — эктодерма (наружный зародышевый листок), б — бластоцель (первичная полость тела) у в — первичная кишка, г — энтодерма (внутренний зародышевый листок), д — бластопор (первичный рот). Зародыш между тем претерпевает сложные превращения. Яйцеклетка начинает дробиться, число клеток эмбриона растет в геометрической прогрессии: 2, 4, 8, 16, 32... Наконец формируется однослойный шар из клеток (бластула). Клетки, находящиеся на одном из полюсов бластулы, мигрируют внутрь, давая начало второму зародышевому листку (энтодерме), из которой позже разовьется кишечник. На этой стадии двуслойный зародыш называется гаструлой. Только на этом этапе у многих животных начинают наконец включаться гены, унаследованные от папы с мамой. У других это происходит чуть раньше — на стадии бластулы. И только млекопитающие — группа, уникальная во многих отношениях, — включают свои гены еще раньше (например, мышь делает это на стадии двух клеток) (Другая уникальная особенность млекопитающих — мощное развитие "геномного импринтинга", механизма, при помощи которого родители целенаправленно влияют на наследственные свойства эмбриона, отчасти — в своих собственных корыстных интересах (см. главу 8). Вполне возможно, что между этими двумя особенностями существует связь. Родители могут быть заинтересованы в том, чтобы "отредактированные" ими гены зародыша включились пораньше). Как удается эмбриону развиваться без всякого "генетического контроля" вплоть до стадии гаструлы? Почему гены зародыша так долго остаются выключенными? Какие механизмы обеспечивают отключение генов в зиготе, а затем их своевременное включение (Последующий текст данного раздела основан на статье: Alexander F. Schier The maternal-zigotic transition: death and birth of RNAs // Science. 2007. v.316. p.406-407.)? Ответ на первый вопрос более или менее ясен. Яйцеклетка содержит большое количество матричных РНК, унаследованных от материнского организма. Эти мРНК считываются с материнских генов заблаговременно, в процессе созревания яйцеклетки. Именно они обеспечивают синтез белков, необходимых для ранних стадий онтогенеза. В определенный момент материнские мРНК начинают уничтожаться. Это происходит как раз тогда, когда зародыш начинает сам производить мРНК, то есть включает свои гены. Этот довольно быстрый процесс замены зародышем материнских мРНК на свои собственные называется maternal- zygotic transition (MZT). Менее ясен вопрос о том, что движет процессом MZT. Предполагается три возможных механизма. 1. По мере роста числа клеток, из которых состоит зародыш, начинает сказываться нехватка тех веществ (что бы они из себя ни представляли), которые не позволяют генам зародыша включиться. Ранние стадии эмбриогенеза животных не случайно называют "дроблением": зигота именно дробится, клетки эмбриона после каждого деления становятся все мельче, поскольку между клеточными делениями отсутствует стадия роста клеток. Общее количество цитоплазмы не растет, тогда как количество клеточных ядер, а следовательно и ДНК, увеличивается в геометрической прогрессии. Если предположить, что яйцеклетка заранее запаслась какими-то ингибиторами транскрипции, то количество этих гипотетических ингибиторов, приходящихся на каждую клетку, должно быстро убывать, и в конце концов их остается так мало, что они уже не могут сдерживать транскрипцию. 2. Не исключено, что в зиготе изначально имеет место целенаправленное блокирование некоторых ключевых генов, работа которых инициирует транскрипцию. Так, показано, что искусственное введение в эмбрион мощных активаторов транскрипции (транскрипционных факторов "широкого спектра действия") может вызвать преждевременное частичное включение эмбрионального генома. 3. Наконец, сама по себе быстрая череда клеточных делений может мешать транскрипции. Ведь каждому делению должно предшествовать удвоение ДНК (репликация). В ходе дробления репликация должна происходить по сути дела непрерывно. Между тем известно, что репликация может мешать транскрипции, а во время клеточного деления (митоза) могут происходить обрыв и уничтожение тех мРНК, синтез которых еще не закончился. Может быть, клетки эмбриона просто физически не успевают транскрибировать свои гены? Искусственное замедление процесса дробления действительно может вызвать преждевременное включение эмбрионального генома. Кстати, у млекопитающих дробление протекает сравнительно медленно, не этим ли объясняется раннее включение генов зародыша? Впрочем, ни одна из этих теорий не объясняет всей совокупности имеющихся фактов. Например, они не могут объяснить, почему эмбриональные гены включаются не все сразу, а постепенно, в строго определенном порядке. Вторым существенным аспектом MZT является уничтожение материнских мРНК. Тут, как выяснилось, все предусмотрено заранее: материнские мРНК помечены особой последовательностью нуклеотидов, расположенной на нетранслируемом (то есть не кодирующем белок) "хвостике" этих молекул. Среди первых генов, которые эмбрион включает в процессе MZT, находятся гены особых белков и маленьких РНК, которые распознают эту последовательность, прикрепляются к ней и тем самым инициируют уничтожение материнских мРНК. До полного понимания всех этих процессов науке еще очень далеко, но дело движется. Когда основные механизмы генной регуляции в ходе раннего онтогенеза будут расшифрованы, биологи смогут вплотную заняться следующим принципиальным вопросом, а именно — зачем все это надо? Почему животные не доверяют своему геному контроль над ранними стадиями развития, а потом в какой-то момент вдруг "переключаются" с материнских мРНК на свои собственные? Между прочим, эта проблема имеет не только теоретическое, но и практическое значение. Трудности, с которыми сталкиваются работы по клонированию животных, во многом определяются тем, что мы еще слишком мало знаем о работе генов в ходе раннего развития. Клонирование животных осуществляется путем пересадки ядра из соматической (неполовой) клетки одного животного в яйцеклетку другого. Однако для нормального развития необходимо, чтобы гены зародыша поначалу помалкивали, а они в донорском ядре вполне активны. Цитоплазма яйцеклетки должна каким-то образом "перепрограммировать" ядро, выключить геном, а потом в нужный момент снова включить его. Пока мы не знаем, как ей в этом помочь, больших успехов в клонировании ожидать трудно. Разгадан механизм регенерации конечностей. "Генетическая программа" индивидуального развития очень тесно связана с другой "программой", на основе которой происходит регенерация — заживление ран и отращивание утраченных частей тела. Изучая регенерацию, ученые одновременно проникают в тайны онтогенеза и наоборот. Биологи из Испании и США недавно провели серию генно-инженерных экспериментов, в результате которых им удалось показать, что регенерация конечностей у позвоночных идет под управлением тех же регуляторных белков, которые руководят развитием конечностей у эмбриона. Белки, о которых идет речь, образуют два сигнально-регуляторных каскада, которые называются Wnt/beta-catenin (о роли белка Wnt в развитии червя Caenorhabditis elegans говорилось выше в сюжете "Развитие червей начинается с хвоста") и BMP (по названию ключевого участника каскада — белка BMP, bone morphogenetic protein). Для первого эксперимента ученые сконструировали специальный вирус, в геном которого был встроен ген белка, блокирующего работу Wnt-каскада. Введение вируса аксолотлю снизило способность к регенерации. У аксолотля в норме отрезанные конечности восстанавливаются полностью, однако у зараженных искусственным вирусом аксолотлей вместо лапы вырастала лишь заостренная культя без пальцев. Этот эксперимент показал, что нормальная работа Wnt-каскада — необходимое условие регенерации, но является ли она также и достаточным условием? Чтобы проверить это, ученые поставили эксперимент на шпорцевой лягушке, у которой в отличие от аксолотля способность к регенерации есть только у головастиков, а с возрастом эта способность теряется. Был сконструирован еще один вирус, производящий белок бета-катенин — один из ключевых участников Wnt-каскада. Введение этого вируса улучшало способность шпорцевых лягушек к регенерации конечностей на тех стадиях развития, когда эта способность уже ослаблена. Однако у взрослых лягушек, полностью потерявших способность к регенерации, этого не наблюдалось. Следовательно, для регенерации необходимо что-то еще, кроме Wnt-каскада.
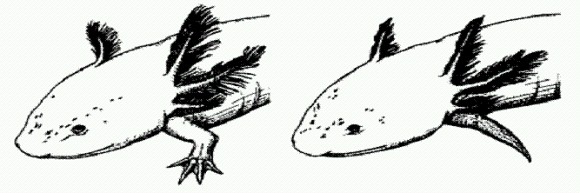
Нормально регенирировавмая после ампутации конечность аксолотля (слева) и недоразвитая конечность у особи, которой был введен Axin1 — ингибитор Wnt-каскада. Третьим подопытным объектом стала рыбка данио (Danio rerio). В норме эта рыбка способна заново отращивать утраченные плавники. Ранее было показано, что различные ингибиторы Wnt-каскада снижают эту способность (так же как у аксолотля и шпорцевой лягушки). Чтобы проверить, способно ли активирование Wnt-каскада усилить способность к регенерации у данио, исследователи использовали рыбок, потерявших эту способность в результате мутации. Введение таким рыбкам бета-катенинового вируса привело к частичному "исправлению" данного дефекта. На данио исследователям удалось также показать, что для успешной регенерации помимо Wnt-каскада должен нормально работать и сигнально-регуляторный каскад BMP. Самое интересное, что оба каскада (Wnt и BMP) руководят и развитием конечностей у эмбрионов. На клеточно-тканевом уровне оба процесса — регенерация и развитие конечности в эмбриогенезе — тоже очень сходны. Все эти данные указывают на то, что восстановление утраченных конечностей осуществляется на основе той же самой программы развития, которая руководит формированием конечностей у эмбриона. И эту программу, по-видимому, можно искусственно "включить" даже у тех животных, которые в норме вообще не способны восстанавливать утраченные конечности. Ведь программа эмбрионального развития есть у всех! Ученым удалось подтвердить это предположение в опытах на курином эмбрионе. Удаление особого многослойного эпителия, расположенного на верхушке зачатка крыла, приводит к недоразвитию крыла. Однако оказалось, что, если активировать у цыпленка ген бета-катенина, крыло после такой ампутации развивается гораздо лучше. Исследователи сделали из всего этого осторожный вывод, что сигнально-регуляторные каскады, управляющие развитием зародыша, вероятно, стали тем "сырым материалом", на основе которого у некоторых животных развилась способность к регенерации. Не исключено, что дальнейшее изучение этих регуляторных систем позволит в будущем "включать" регенерацию у взрослых животных, полностью лишенных способности к восстановлению утраченных частей тела, в том числе и у человека. (Источник: Yasuhiko Kawakami et al. Wnt/beta-catenin signaling regulates vertebrate limb regeneration // Genes & Development. 2006. V. 20. P. 3232-3237.) Кембрийский взрывВ самом начале кембрийского периода, примерно 542 млн лет назад, многие группы животных почти одновременно стали обзаводиться твердым минерализованным скелетом. Поскольку в ископаемом состоянии обычно сохраняются именно такие скелеты, а мягкие части бесследно исчезают, это событие в палеонтологической летописи выглядит как внезапное, "взрывное" появление многих групп животных (моллюсков, членистоногих, губок, археоциат, брахиопод, к которым несколько позже присоединяются иглокожие, кораллы, мшанки и другие). Отсюда и общепринятое название этого события — "кембрийский взрыв". Вся та палеонтология, о которой мы говорили до сих пор, — палеонтология докембрия, то есть изучающая архейский и протерозойский эоны со всеми биомаркерами, окремненными цианобактериями, акритархами, городискиями и мягкотелыми животными венда, — стала интенсивно развиваться лишь сравнительно недавно. До этого момента докембрийские толщи казались ученым практически мертвыми, не содержащими почти никаких следов жизни. "Кембрийский взрыв" выглядел внезапным появлением множества разнообразных организмов словно бы из ниоткуда. Поэтому докембрий назвали криптозоем — временем "скрытой жизни", а последний этап развития биосферы, начавшийся с кембрия и включающий палеозойскую, мезозойскую и кайнозойскую эры, носит название фанерозоя (время "явной жизни"). Дарвин считал кембрийский взрыв одним из фактов, не укладывающихся в его теорию постепенных эволюционных изменений. Впоследствии выяснилось, что "взрыв" на самом деле был не таким уж взрывообразным. Как мы теперь знаем, предки многих кембрийских групп жили и раньше, но они были по большей части бесскелетными, мягкотелыми. Именно поэтому палеонтологи долго не могли обнаружить их остатков в докембрийских породах. Загадка кембрийского взрыва, тем не менее, осталась, только теперь речь идет не о внезапном возникновении как бы "из ничего" многих типов животных, а о более или менее одновременном появлении у них минерального скелета. Это могло быть связано с изменениями условий среды. Например, к такому эффекту могло бы привести резкое уменьшение кислотности воды, в результате чего карбонат кальция (СаСО3) — самый распространенный у животных скелетообразующий материал — стал хуже растворяться в морской воде и легче выпадать в осадок. Предложен и целый ряд других объяснений. Хорошие популярные рассказы о кембрийском взрыве и теориях, предложенных для его объяснения, можно найти в книгах А. Ю. Розанова "Что произошло 600 миллионов лет назад" (1986) http://www.lib.prometey. org/?id=i4599 и К. Ю. Еськова "История Земли и жизни на ней" (2000) http://warrax.net/51/eskov/cover_eskov.htm. Обе книги, к счастью, находятся в открытом доступе в Интернете, поэтому повторяться нет необходимости. Я расскажу лишь об одном недавнем исследовании, которое имеет отношение одновременно и к кембрийскому взрыву, и к сравнительной геномике, и к ранней эволюции животных, и к некоторым идеям, изложенным выше в этой главе. Для того чтобы дружно обзавестись карбонатными (то есть сделанными из СаСО3) скелетами, животным недостаточно было одних лишь благоприятных условий среды. Нужны были еще специальные гены и ферменты, при помощи которых животные могли бы контролировать образование и рост кристаллов карбоната кальция в нужных местах и в нужном количестве. Важнейшую роль в образовании карбонатных скелетов у животных играют ферменты карбоангидразы, которые примерно в миллион раз ускоряют реакцию превращения растворенного в воде углекислого газа в гидрокарбонат: C02 + H20-> carbonyc anhydrase HCO3- + H+ Карбоангидразы широко распространены в живом мире, в том числе и у прокариот. Помимо биоминерализации они участвуют в выполнении множества других функций (регуляция рН, транспорт ионов, выведение С02 из тканей и др.). До сих пор оставалось неясным, когда и в какой последовательности разные группы животных обзаводились карбоангидразами. Возможно, они уже имели эти ферменты к началу "кембрийской скелетной революции" — в этом случае надо признать, что скелетообразование не является первичной функцией карбоангидраз у животных, но могло быть и так, что независимое приобретение генов карбоангидраз разными группами организмов как раз и послужило толчком к появлению скелетов. Для прояснения ситуации очень не хватало геномных данных по самым примитивным многоклеточным животным, таким как губки. Биологи из Германии и Австралии недавно восполнили этот пробел, изучив скелетообразующие ферменты у примитивной архаичной губки Astrosclera willeyana (Daniel J. Jackson et al. Sponge paleogenomics reveals an ancient role for carbonic anhydrase in skeletogenesis // Science. 2007. v.316. p. 1893-1895.) Это настоящее "живое ископаемое": род Astrosclera существует больше 200 млн лет (с конца триасового периода), а по строению своего карбонатного скелета эта губка очень близка к формам, процветавшим еще в палеозое (так называемым строматопоратам). Скелет астросклеры состоит из мелких сферических элементов, которые постепенно растут и сливаются друг с другом. Ученые выделили из скелета губки органическую фракцию, а из нее — все белки. Три преобладающих белка оказались карбоангидразами. Исследователи определили их аминокислотную последовательность, а затем по этой последовательности "выудили" из генома и три соответствующих гена. Это позволило, сравнивая между собой нуклеотидные последовательности генов карбоангидраз примитивной губки и высших животных, чьи геномы уже прочтены, реконструировать эволюцию этих белков у животных. Ученые пришли к выводу, что все многочисленные и разнообразные карбоангидразы животных происходят от одного предкового белка, который имелся у последнего общего предка всех животных. В разных эволюционных линиях ген этой исходной карбоангидразы неоднократно подвергался независимым дупликациям (удвоениям). Так возникали различные новые варианты карбоангидраз. "Последний общий предок всех животных", вне всяких сомнений, жил задолго до кембрийской скелетной революции. Получается, что животные изначально были хорошо подготовлены (преадаптированы) к развитию минерального скелета — у них с самого начала были ферменты, способные резко ускорить образование карбоната кальция. Эти ферменты, очевидно, использовались докембрийскими мягкотелыми животными для других целей — как уже говорилось, у карбоангидраз в животном организме хватает работы и без скелетообразования. Когда условия среды стали благоприятствовать биоминерализации, разные группы животных не сговариваясь "привлекли" часть своих карбоангидраз к выполнению новой функции. Совсем недавно была опубликована еще одна интересная работа, посвященная исследованию геномов губок (Onur Sakarya et al. A Post-Synaptic Scaffold at the Origin of the Animal Kingdom // PLoS One. 2007. 2(6): e506). Оказалось, что у этих примитивных многоклеточных уже есть значительная часть комплекса так называемых постсинаптических белков, которые у более высокоорганизованных животных функционируют в нервных клетках и участвуют в "приеме сигнала". У губок, однако, нет нервных клеток. Зачем же им эти белки? По всей видимости, они участвуют в обмене сигналами между клетками губки. Животное может не иметь нервной системы, но если его клетки совсем не будут "общаться" друг с другом, это будет уже не животное, а скопление одноклеточных организмов. Позже, когда у животных развилась нервная система, эти "коммуникационные" белки пригодились для формирования системы обмена сигналами между нервными клетками. Этот пример, как и множество других, показывает, что большинство эволюционных новшеств возникает не на пустом месте, а собирается из "подручного материала", причем часто для радикального изменения функции какого-нибудь белка или белкового комплекса достаточно совсем небольших генетических изменений. Что почитать на эту тему в ИнтернетеК. Ю. ЕСЬКОВ. История Земли и жизни на ней. 2000. http://warrax.net/51/eskov/cover_eskov.html A. Ю. ЖУРАВЛЕВ. Скелетный докембрий. 2006. http://vivovoco.rsl.ru/vv/journal/nature/12_06/paleo.htm B. В. МАЛАХОВ. Происхождение билатерально-симметричных животных. 2004. http://evolbiol.ru/malahov.htm В. В. МАЛАХОВ. Происхождение хордовых животных. 1996. http://evolbiol.ru/malahov1996.pdf Я. Е. МАЛАХОВСКАЯ, А. Ю. ИВАНЦОВ. Вендские жители земли. 2003. http://evolbiol.ru/vend.htm А. Ю. РОЗАНОВ. Что произошло 600 миллионов лет назад. 1986. http://www.lib.prometey.org/?id=i4599 М. А. ФЕДОНКИН. Две летописи жизни: опыт сопоставления (палеобиология и геномика о ранних этапах эволюции биосферы). 2006. http://evolbiol.ru/fedonkin2006.htm И. И. ШМАЛЬГАУЗЕН. Организм как целое в индивидуальном и историческом развитии. 1982. http://rogov.zwz.ru/Macroevolution/org_kak_tsel.doc |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Вверх |
||||
|
|
||||