|
||||
|
|
ГЛАВА 3. ВЕЛИКИЙ СИМБИОЗЭволюция "с человеческим лицом"Типичным примером примитивизации дарвиновских идей является широко распространенное по сей день мнение о том, что в основе эволюции лежат исключительно эгоизм, конкуренция и безжалостная "борьба за существование". Каждый за себя, кто смел — тот и съел, побеждает сильнейший... Одним словом, сплошной "закон джунглей". Сколько раз недобросовестные публицисты — и особенно креационисты — ставили биологам в вину эти взгляды, утверждая, что дарвинизм якобы подвел научную базу под самые несправедливые и бесчеловечные политические системы, такие как фашизм или, скажем, колониализм. Забывая при этом, что дело ученых (по крайней мере, ученых-естественников, с гуманитариями ситуация сложнее) — искать истину, а не решать, что хорошо, а что плохо. Даже если бы эволюция и впрямь основывалась исключительно на "праве сильного" (что, как мы видели в предыдущей главе, далеко не всегда так), из этого вовсе не следует, что на таком же принципе должны строиться законы человеческого общества. В конце концов, не следует же из закона всемирного тяготения, что мы все должны лежать пластом, никогда не пытаясь встать! Конкуренция, конечно, играет весьма важную роль и в биологической, и в социальной эволюции. Однако в конечном счете, как правило, в выигрыше оказываются не те, кто сумел добиться безраздельного господства в той или иной сфере и уничтожить всех конкурентов, а те, кому удалось наладить взаимовыгодное сотрудничество с ними и превратить врагов в друзей. Если уж на то пошло, для социальной эволюции "закон джунглей" характерен даже в большей степени, чем для эволюции биологической. История полна примеров безжалостного истребления и вытеснения одними народами других, и этническое разнообразие человечества в целом скорее снижается, чем растет (особенно это заметно в наши дни из-за так называемой глобализации). Для биологической эволюции такие явления менее характерны. Новые "прогрессивные" группы обычно не вытесняют старые, а добавляются к ним, что ведет к неуклонному росту биологического разнообразия. Кстати, рост биологического разнообразия, как выяснилось, хорошо описывается гиперболической функцией, точно так же, как и рост численности (но не разнообразия!) человечества. Например, появление многоклеточных организмов вовсе не привело к вытеснению одноклеточных. Этого и не могло произойти, потому что высшие организмы изначально развивались в среде, населенной низшими; эти последние составляли важнейшую часть тех "внешних условий", к которым должны были адаптироваться высшие. В результате низшие организмы и по сей день остаются жизненно необходимы для существования высших. Если сегодня на Земле вдруг исчезнут все одноклеточные, биосфера испытает немедленный коллапс. Растительность придет в упадок, потому что никто на свете, кроме некоторых микроорганизмов, не умеет фиксировать атмосферный азот (см. главу "Планета микробов"). А если бы даже каким-то чудом уцелели растения, это не спасло бы животных, ведь они не способны переваривать самые массовые виды растительной пищи без помощи кишечных микроорганизмов. Если же исчезнут многоклеточные, на процветании мира микробов это отразится в гораздо меньшей степени. Без сотрудничества, кооперации, симбиоза не может существовать (и тем более развиваться) ни одна живая система. Даже для самых жестоких человеческих коллективов (изолированных, так называемых карцерных, где всплывают на поверхность самые дремучие инстинкты и "архаическое мышление") не характерны взаимоотношения по принципу "каждый против каждого". Неизбежно будут складываться какие-то группировки, союзы, альянсы.
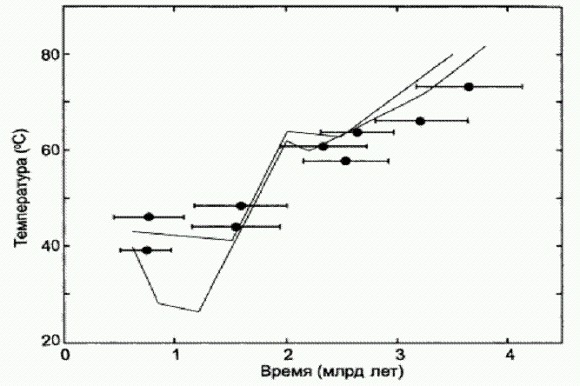
Рост разнообразия семейств морских (слева) и континентальных (справа) животных и растений в течение фанерозойского зона (последние 542 млн лет истории Земли). По горизонтальной оси — время в млн лет назад. Толстой линией показан гиперболический тренд. Из статьи: Марков А. В., Коротаев А. В. Гиперболический рост разнообразия морской и континентальной биот фанерозоя и эволюция сообществ. 2008. //Журнал общей биологии. 2008. № 3. В биологии необходимость кооперации и симбиоза совершенно очевидна. Для того чтобы выжить и оставить потомство, каждое живое существо должно справиться с множеством разнообразных проблем. Нужно каким-то образом получать из окружающей среды необходимые вещества, а недостающие самостоятельно синтезировать из подручного материала; нужно добывать энергию, необходимую для энергоемких химических и физических процессов; нужно вовремя избавляться от отходов жизнедеятельности, находить подходящих партнеров для обмена наследственным материалом, заботиться о потомстве, защищаться от хищников и так далее — и все это в переменчивой, далеко не всегда благоприятной внешней среде. Требования, предъявляемые жизнью к каждому отдельному организму, не только многочисленны и разнообразны — очень часто они еще и противоречивы. Невозможно оптимизировать сложную систему сразу по всем параметрам: чтобы добиться совершенства в чем-то одном, приходится жертвовать другим. Поэтому эволюция — это вечный поиск компромисса, и отсюда следует неизбежная ограниченность возможностей любого отдельно взятого живого существа. Самый простой и эффективный путь преодоления этой ограниченности — симбиоз, то есть кооперация "специалистов разного профиля". Биологическая эволюция в целом производит впечатление гораздо более "гуманного" процесса, чем история заселения европейцами Нового Света или процесс формирования карцерных коллективов. Как мы увидели из предыдущих глав, симбиоз и кооперация стали неотъемлемыми свойствами земной жизни с самого момента ее зарождения, и в дальнейшем эти тенденции только усиливались. Примерно 2 млрд лет назад они привели к радикальному перелому в развитии жизни на нашей планете — к событию, которое по праву считается вторым по значимости в ее истории. На первом месте, конечно, стоит само появление жизни, о котором мы говорили в главе 1. А появление эукариотической (ядерной) клетки уверенно занимает второе место (Многие, в том числе и я, считают, что на третье место надо поставить появление человека разумного. Хотя не исключено, что здесь сказывается наш неистребимый антропоцентризм). Сообщество превращается в организм"Планета микробов" жила и успешно развивалась в течение миллиарда или более лет, и за это время ее микроскопические обитатели добились немалых успехов. Высшим достижением этого этапа эволюции стали сложные микробные сообщества — бактериальные маты, о которых говорилось в предыдущей главе. По уровню целостности бактериальный мат приближается к настоящему организму. Приближается, но все-таки не достигает этого уровня. В какой-то момент эволюция микробных сообществ словно наткнулась на невидимую преграду — и вдруг оказалось, что для дальнейшего эволюционного прогресса не хватает чего-то очень важного. Забегая вперед, скажу, что прокариоты так и не смогли преодолеть этот рубеж и дать начало настоящим многоклеточным организмам. Все развитие прокариотной биосферы строилось на основе одного и того же базового "кирпичика", породившего бесчисленное множество собственных модификаций. Этим кирпичиком была прокариотическая клетка. При всех бесспорных преимуществах этой универсальной живой единицы, при всем ее умении приспосабливаться почти к любым условиям и извлекать энергию чуть ли не из любого химического процесса в строении прокариотической клетки есть ряд непреодолимых ограничений. Главное из них связано с отсутствием клеточного ядра. Наследственный материал прокариот (обычно это единственная кольцевая молекула ДНК — кольцевая хромосома) находится прямо в цитоплазме, то есть, образно говоря, в бурлящем биохимическом котле, где происходит обмен веществ и осуществляются тысячи химических реакций. В такой неспокойной обстановке очень трудно развить сложные и эффективные молекулярные механизмы регуляции работы генов. У эукариот функционирование генома регулируется сотнями и тысячами специализированных белков, а также особыми регуляторными РНК и другими молекулами. Весь этот управляющий аппарат находится в ядре клетки, и ядерная оболочка надежно защищает его от бурной биохимической деятельности цитоплазмы. Тонкая регуляция работы генов обеспечила эукариотам качественно иной уровень пластичности. Самое главное, она позволила клетке радикально менять свои свойства, структуру и облик, не изменяя при этом сам геном, а только усиливая или ослабляя работу разных генов. Именно эта пластичность позволила эукариотам в конце концов стать многоклеточными в строгом смысле этого слова. Ведь в настоящем многоклеточном организме не просто много клеток, а много разных типов клеток (покровные, мышечные, нервные, половые и т. д.) Однако геном у них у всех один и тот же! Разумеется, механизмы регуляции работы генов есть и у прокариот, но они проще и работают менее эффективно. Некоторые "высшие" прокариоты, такие как цианобактерии, могут даже иногда менять строение своих клеток, не меняя генома (пример — образование гетероцист у нитчатых цианобактерий, о чем говорилось в главе "Планета микробов"). Но эти зачаточные способности к адаптивным модификациям клеток не идут ни в какое сравнение с тем, что наблюдается у эукариот. Второй важный конструктивный "недостаток" прокариотической клетки состоит в отсутствии окруженных мембранами органелл. Или, как говорят биологи, в отсутствии компартментализации внутренней среды клетки: цитоплазма прокариот не подразделена на отсеки — "компартменты". А ведь далеко не все биохимические процессы, которые могли бы оказаться полезными клетке, можно осуществлять в едином "общем котле", каковым является цитоплазма прокариот. Представьте себе, как усложнилась бы работа химика, если бы в его распоряжении имелась лишь одна-единственная пробирка! Прокариоты, конечно, пытались по-своему преодолеть этот недостаток. Если присмотреться к прокариотической клетке повнимательнее, можно заметить, что в распоряжении бактерий на самом деле не одна "пробирка", а две. В роли второй выступает так называемое периплазматическое пространство, то есть область снаружи от клеточной мембраны (здесь обычно расположено толстое, рыхлое, сложно устроенное многослойное образование — "клеточная стенка"). В толще клеточной стенки могут протекать химические процессы, не совместимые с теми, которые идут в цитоплазме. Но и двух пробирок все-таки мало для хорошей химической лаборатории! Эукариоты в этом отношении дадут прокариотам сто очков вперед. Внутренняя среда эукариотической клетки подразделена двойными и одинарными мембранами на множество разнообразных отсеков-"компартментов" (ядро, митохондрии, пластиды, эндоплазматическая сеть и т. д.). Именно из-за этих ограничений прокариоты с самого начала так сильно тяготели к симбиозу. В микробном сообществе разные виды микробов в функциональном отношении соответствуют разным отсекам эукариотической клетки. Сообща микробы могут достичь гораздо большего, чем поодиночке. Однако как бы ни были тесны связи в симбиотическом содружестве микробов, сообщество не становится настоящим целостным организмом, потому что оно не может размножаться как единое целое (подробнее об этом см. врезку "Появлению многоклеточности мешают обманщики" в главе "Рождение сложности", стр...). Каждый из микробов сохраняет способность "сбежать" из сообщества и перейти к самостоятельной жизни, если где-то сложатся подходящие для этого условия; каждый размножается сам по себе и имеет свой собственный индивидуальный геном, который и передает своим потомкам. Поэтому естественный отбор продолжает действовать на уровне отдельных клеток, а не на уровне сообщества. Любая мутация, повышающая жизнеспособность данного конкретного микроба, будет поддержана отбором даже в том случае, если это вредно для сообщества как целого. Чаще всего вред для сообщества будет означать и вред для данного микроба, но, увы, не всегда — иначе не было бы на свете паразитов и обманщиков. Поэтому эволюция прокариот в основе своей остается эгоистической. Чтобы преодолеть эти ограничения, прокариотическим клеткам нужно было сделать еще один шаг — вполне естественный и логичный — в сторону дальнейшего усиления интеграции, сплоченности сообщества. Они должны были по-настоящему слиться в единый организм, отказаться от своей клеточной индивидуальности и объединить свои персональные хромосомы в один большой общий геном. Именно это и случилось в начале протерозойского эона (вероятно, около 2,0-2,2 млрд лет назад) (в 1999 году в отложениях возрастом 2,7 млрд лет были найдены следы присутствия эукариот - фрагменты органических молекул (мембранных стеролов), которые ни у кого, кроме эукариот, не встречаются. Поэтому до самого недавнего времени многие эксперты считали, что эукариоты появились еще в конце архейской эры. Однако в октябре 2008 года ученые, открывшие древние стеролы, сами же и "закрыли" свое открытие. Они установили, что стеролы просочились в древние породы из вышележащих, более молодых слоев). Сообщество прокариот, слившееся в единый организм — эукариотическую клетку, — стало новым базовым "строительным блоком", усовершенствованным "кирпичиком" в том великом конструкторе, из которого эволюция по сей день продолжает собирать новые формы жизни. Воскрешенные белки рассказывают о климате древнейших эпох. Для реконструкции древнейших этапов эволюции очень важно хотя бы примерно представлять, в каких условиях происходили эти события. Одним из самых спорных является вопрос о том, каким был климат на планете в течение архейского и протерозойского эонов. Как правило, о климате столь отдаленных эпох судят по изотопному составу углерода, кислорода, кремния, серы и других элементов в осадочных породах земной коры. Эти данные порой допускают неоднозначное толкование. Иногда предположения о протерозойских температурах базируются на присутствии в породах ископаемых остатков тех или иных бактерий, но и эти данные считаются не вполне надежными и обычно вызывают массу возражений. Как и в случае с абсолютными геохронологическими датировками (Популярный обзор современных методов геохронологии см. в статье А.В. Марков Хронология далекого прошлого http://elementy.ru/lib/430055), для повышения надежности и точности палеоклиматических реконструкций огромное значение имеет привлечение разных источников данных. Если каждый метод в отдельности не очень надежен, нужно использовать сразу несколько независимых методов и смотреть, сходятся ли результаты. Вплоть до самого последнего времени палеоклиматические реконструкции, относящиеся к архею и протерозою, казались весьма сомнительными. Однако в феврале 2008 года в журнале Nature появилась статья американских исследователей, которая сильно изменила ситуацию к лучшему. Ученые из Фонда прикладной молекулярной эволюции, факультета биохимии и молекулярной биологии Флоридского университета и компании "ДНК 2.0м (DNA 2.0 lnс) провели многоступенчатое исследование, в результате которого удалось получить весьма правдоподобную реконструкцию температуры земной поверхности в архейские и протерозойские времена, то есть 3,5-0,5 млрд лет назад. Ученые подошли к проблеме с совершенно новой и неожиданной стороны. Их идея состояла в том, чтобы восстановить белки древнейших бактерий и проверить, к каким температурам эти белки лучше всего приспособлены — благо современная молекулярная биология уже способна выполнить такие удивительные маневры, как реконструирование исчезнувших белков. Тогда диапазон температур, в которых реконструированные белки будут устойчивы, как раз и покажет температурные условия, к которым были приспособлены бактерии. В качестве белков-индикаторов решено было восстановить ферменты, участвующие в последовательном присоединении аминокислот к синтезируемой молекуле белка (этот процесс называется элонгацией и представляет собой основной этап трансляции). Ферменты этого этапа трансляции называются факторами элонгации. Они должны всегда присутствовать в клетке в большом количестве, пока клетка жива, и всегда должны быть наилучшим образом приспособлены к окружающей среде, иначе белки в клетке будут синтезироваться слишком медленно. Исследователи остановили свой выбор на одном из трех основных факторов элонгации, присутствующих у прокариот, — EF-Tu (elongation factor thermo-unstable). Исходя из строения факторов элонгации и, соответственно, кодирующих их генов у разных групп современных бактерий, а также у хлоропластов и митохондрий были реконструированы наиболее вероятные предковые гены факторов элонгации, соответствующие основным узлам (точкам ветвления) эволюционного древа бактерий. Эта стандартная статистическая задача сейчас легко решается: банки данных с расшифрованными генными последовательностями, в том числе и гены белков — факторов элонгации, находятся в открытом доступе. Для реконструкций были использованы две альтернативные схемы эволюции бактерий, так что генные последовательности пред- ковых белков получились немного разные. Затем эти гипотетические белки изготовили в лаборатории: собрали кодирующие их гены, внедрили их в клетки кишечной палочки, и бактерия вынужденно синтезировала необходимые ученым белки. После чего можно было спокойно изучать термостойкость этих воскрешенных белков. Выяснилось, что, чем раньше разошлись эволюционные ветви, то есть чем раньше жил общий предок соответствующей группы бактерий, тем более термостойким был его фактор элонгации. Самые древние общие предки, жившие в раннеархейские времена (3,5 млрд лет назад), были приспособлены к температуре около 60-70°С. Самые молодые, жившие в конце протерозоя (550 млн лет назад), предпочитали гораздо более прохладный климат — 37-35°С.
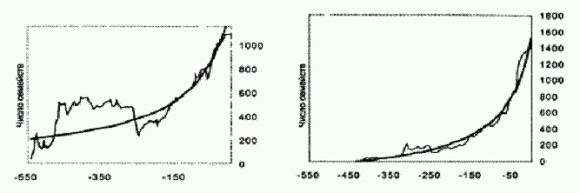
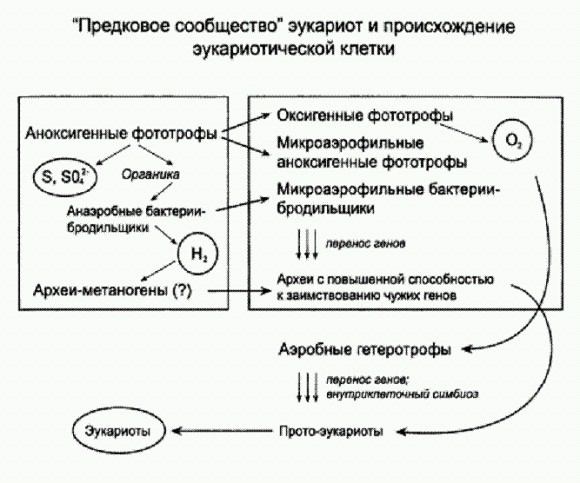
Снижение воссозданных температур плавления факторов элонгации по мере приближения к современности. По горизонтальной оси — время в млрд лет назад. Горизонтальные отрезки — это предполагаемое время существования общего предка той или иной группы бактерий по молекулярным часам с диапазоном возможных ошибок. Температура плавления экспериментальных белков определена без ошибок (для современных бактерий показано, что они лучше всего растут при температуре примерно на 2 градуса ниже у чем температура плавления белка EF-Tu, характерного для данной бактерии). Серыми линиями показан тренд снижения температуры океанов по изотопам кислорода (светлая и темная линии отражают оценки, полученные разными авторами на основе анализа разных горных пород). Это означает, что на заре земной жизни бактерии жили примерно в таких же условиях, какие сейчас существуют в горячих источниках, если к этому прибавить ультрафиолет и отнять кислород (пока в атмосфере не было кислорода, не было и озонового слоя, задерживающего ультрафиолетовое излучение). Что касается цианобактерий, то они, как выяснилось, изначально жили при температуре около 64°С. Примерно к таким же температурам приспособлены и современные цианоактериальные маты, живущие в горячих источниках. Общий предок всех митохондрий, судя по свойствам воскрешенных белков, жил при температуре 51-53°С; общий предок всех бактерий — вообще при 64,8-73,3°С. Самое главное, что полученные результаты почти полностью совпали с теми графиками, которые были получены ранее по изотопам кислорода и кремния. Совпадение результатов, полученных различными методами и на основе различных данных, всегда обнадеживает — это, пожалуй, единственный критерий правдоподобия подобных реконструкций. (Источник: Eric A. Gaucher, Sridhar Govindarajan, Omjoy К. Ganesh. Palaeotemperature trend for Precambrian life inferred from resurrected proteins // Nature. 2008. V. 451. P. 704-707.) Из кусочковВ настоящее время в научной литературе обсуждается два или три десятка возможных сценариев превращения сообщества прокариот в эукариотическую клетку Они разнятся в деталях, но имеют много общего в целом. Общепризнанными считаются следующие факты. 1. Митохондрии (органеллы эукариотической клетки, ответственные за кислородное дыхание) являются прямыми потомками прокариот из группы альфапротеобактерий. К этой группе бактерий относятся современные внутриклеточные паразиты риккетсии (такие как возбудитель сыпного тифа, а также вольбахия, о которой рассказано в главе "Наследуются ли приобретенные признаки?"), некоторые аноксигенные фототрофы — пурпурные бактерии — и многие другие микробы. В качестве неоспоримых доказательств своего бактериального происхождения митохондрии по сей день сохранили маленькую кольцевую хромосому прокариотического типа с несколькими функционирующими генами (все остальные ми- тохондриальные гены перебазировались в ядро и стали частью "общего" ядерного генома клетки), способность к самостоятельному размножению путем деления (причем осуществляется это деление при помощи тех же самых белков, что и у свободноживущих бактерий), типично бактериальный аппарат синтеза белка и множество других признаков. Сомневаться не приходится: предки митохондрий, свободноживущие альфапротеобактерии, когда-то были "проглочены" другим одноклеточным организмом, но не переварились, а стали неотъемлемой частью клетки. 2. Пластиды (органеллы растительной клетки, отвечающие за фотосинтез) являются потомками цианобактерий. Сходство пластид со свободноживущими цианобактериями настолько велико, что никаких сомнений тут просто не может быть. 3. Что касается "всего остального", то есть цитоплазмы эукариотической клетки и ее ядра, то здесь наблюдается причудливое смешение признаков архей и некоторых групп бактерий, а также уникальных черт, свойственных только эукариотам. Основные различия между имеющимися гипотезами происхождения эукариот касаются происхождения "ядерно-цитоплазматического компонента" эукариотической клетки. Ясно, что его предком был какой-то крупный одноклеточный организм, который "проглотил" сначала будущих митохондрий, а потом будущих пластид, и превратил их в своих внутренних "сожителей"-симбионтов. Или, может быть, они не были проглочены, а проникли в него по собственной инициативе (подобно тому, как сейчас это делают внутриклеточные паразитические бактерии). Проблема в том, что этот организм-хозяин, насколько мы можем судить, был не очень похож на современных, доживших до наших дней прокариот. Он обладал рядом уникальных свойств. Существует несколько версий его происхождения. Одни эксперты считают, что это была архея, возможно, близкая к современным термоплазмам или ферроплазмам, о которых мы немного рассказали в главе "Происхождение жизни". Все уникальные свойства развились уже после приобретения внутренних симбионтов (митохондрий) и объединения разнородных геномов в единый ядерный геном. Согласно другой гипотезе, предком цитоплазмы и ядра эукариот был представитель не архей и не бактерий, а некоей особой вымершей группы прокариот. Согласно третьей точке зрения, это был химерный организм, образовавшийся в результате слияния клеток нескольких разных архей и бактерий. Впрочем, дело могло обойтись и без слияния — оказалось достаточно очень интенсивного обмена генами между разными микробами, чтобы их свойства перемешались в одной клетке (о горизонтальном генетическом обмене подробно рассказано в главе "Наследуются ли приобретенные признаки?").
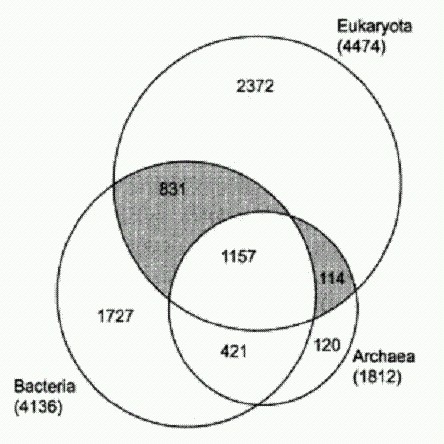
Распределение белковых семейств в трех надцарствах живой природы. Площади кругов соответствуют количеству семейств белков, встреченных у представителей данного надцарства. Как видно из рисунка, 1157 белковых семейств являются общими для всех трех надцарств, 2372 встречены только у эукариот, 831 есть у эукариот и бактерий, но не у архей, и т. д. (из статьи: Марков А. В., Куликов А. М. Происхождение эвкариот: выводы из анализа белковых гомологий в трех надцарствах живой природы // Палеонтол. журн. 2005. № 4. С. 3-18. http://evolbiol.ru/markov_kulikov.htm). У каждой из версий, понятное дело, есть свои аргументы и свои сторонники. Лично мне больше всего нравится "химерная" теория. По крайней мере, именно к ней склонил нас с А. М. Куликовым (Институт биологии развития РАН) сравнительный анализ семейств белков, имеющихся у представителей трех надцарств живой природы — архей, бактерий и эукариот. Белки архейного происхождения, хотя их не очень много (114 семейств, см. рисунок), играют в эукариотической клетке ключевую роль. Именно они отвечают за работу с генетической информацией — транскрипцию, трансляцию, репликацию. Сюда относятся и НК-полимеразы, о которых шла речь выше, а также большинство белков, входящих в состав рибосом. Это позволяет предполагать, что в основе ядерно-цитоплазматического компонента эукариот лежала именно архея, а не бактерия и не какая-то особая "третья" группа прокариот, не дожившая до наших дней. Многие белки бактериального происхождения попали в эукариотическую клетку вместе с "проглоченными" симбионтами — предками митохондрий и пластид. Однако среди "бактериальных" белков цитоплазмы и ядра есть и много таких, которые, скорее всего, не могли быть получены таким способом. Речь идет о тех семействах белков, которые есть у эукариот и есть также у бактерий, но не у тех, от которых произошли органеллы, а у каких-нибудь других. Иными словами, в эукариотической клетке существуют "бактериальные" семейства белков, которые не могли быть получены первыми эукариотами ни от "проглоченных" альфапротеобактерий, ни от цианобактерий — предков пластид. Но они, однако, могли быть получены от других бактерий — в особенности от различных бродильщиков (гетеротрофных бактерий, сбраживающих углеводы в бескислородных условиях). Похоже, именно от бродилыциков эукариоты получили, в частности, ферменты гликолиза — так называется важнейший энергетический процесс, происходящий в цитоплазме эукариотической клетки. Суть его в том, что молекула глюкозы расщепляется ("сбраживается") без использования кислорода до пировиноградной кислоты (пирувата), и при этом происходит синтез АТФ. Пируват является для цитоплазмы конечным продуктом обмена, "отходом жизнедеятельности". Но пируват затем попадает в митохондрии, которые "сжигают" его в своей кислородной печке с огромным выходом энергии (которая тоже используется для синтеза АТФ). В совокупности бескислородный гликолиз, происходящий в цитоплазме, и кислородное дыхание, происходящее в митохондриях, являются главными источниками АТФ для эукариотической клетки.
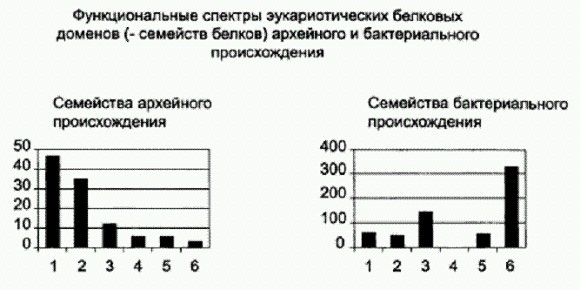
Распределение эукариотических белков архейного и бактериального происхождения по шести функциональным группам: 1 — синтез белка, 2 — репликация, транскрипция, модификация ДНК и РНК, 3 — сигнальные и регуляторные белки, 4 — образование мембранных пузырьков, 5 — транспортные и сортировочные белки, 6 — обмен веществ. И вот на основе всех этих данных, а также с учетом того, что известно о строении и функционировании микробных сообществ, вырисовывается следующий возможный сценарий превращения сообщества прокариот в эукариотическую клетку (конечно, надо помнить, что это только один из возможных сценариев). Этапы великой драмыВ начале, как мы уже знаем, было сообщество. Это был трехслойный бактериальный мат, почти такой же, как современные бактериальные маты, с той разницей, что верхний его слой образовывали не кислородные (оксигенные), а бескислородные фотосинтетики. Это были предки цианобактерий, еще не научившиеся использовать в качестве донора электрона воду. Они по старинке потребляли сероводород и выделяли серу или сульфаты. Второй слой составляли другие аноксигенные фотосинтетики, в том числе альфапротеобактерии — предки нынешних пурпурных бактерий (а заодно и митохондрий, но об этом чуть позже). Эти розовые создания и сегодня живут в бактериальных матах под слоем цианобактерий, потому что питаются более длинноволновым светом, который легко проходит сквозь верхний зеленый слой сообщества. В третьем слое жило много всякой мелочи. Здесь были бактерии-бродильщики, которые сбраживали избыточную органику, производимую верхними фотосинтезирующими слоями. Они выделяли молекулярный водород, который использовался для восстановления сульфатов бактериями-сульфатредукторами. В результате их деятельности в сообществе пополнялись запасы сероводорода, необходимого двум верхним слоям. Здесь же подвизались и археи-метаногены, которые с удовольствием использовали производимый бродильщиками водород для восстановления углекислого газа и синтеза метана. Метаногены и сегодня живут практически везде, где нет кислорода и есть бродильщики, — например, у нас в кишечнике.
Сообщество было вполне устойчивым и могло спокойно существовать в таком виде сотни миллионов лет (что оно, по всей видимости, и делало). Но потом цианобактерии "изобрели" кислородный фотосинтез (см. предыдущую главу), начали выделять кислород, и спокойному существованию пришел конец. Для всех древних форм земной жизни — и для всех без исключения членов нашего древнего сообщества — кислород был опаснейшим ядом. Даже самим цианобактериям было не очень приятно жить в отравленной — с их точки зрения — среде. Но возможность наконец-то избавиться от "сероводородной зависимости" перевешивала все прочие соображения. Конечно, цианобактерии поступили крайне эгоистично — ради собственной независимости они чуть не отравили все живое на планете, но в конечном счете их эгоизм оказался полезен для биосферы. Ведь без него наша Земля и по сей день оставалась бы "планетой микробов". К счастью для цианобактерий, они очень быстро нашли способ обезвреживать ядовитые продукты собственной жизнедеятельности. И тот же самый способ — причем даже с большей эффективностью — применили для защиты от яда обитатели второго слоя, пурпурные бактерии. Скорее всего, тут не обошлось без горизонтального обмена генами. В чем же состоял этот способ? Как обычно, эволюция слепила новую молекулярную "машинку" из того, что первым подвернулось. Для эволюции это очень характерный и легко узнаваемый стиль. Поскольку в данном случае дело касалось фотосинтезирующих микробов, в ход пошел аппарат фотосинтеза. Небольшая модификация некоторых частей этого аппарата привела к возникновению системы кислородного дыхания. Как клетки научились дышать. Упрощенно говоря, в процессе фотосинтеза квант света выбивает из молекулы хлорофилла электрон. Этот "возбужденный" электрон затем передается "из рук в руки" по цепочке белков, постепенно теряя свою энергию, которая идет на синтез АТФ. В конце концов электрон возвращается на место, то есть передается молекуле хлорофилла — той же самой или другой. "Генеральная идея" кислородного дыхания состоит в том, что электрон берется не у хлорофилла, а у какой-нибудь другой органической молекулы (например, у пирувата — уже упоминавшейся пировиноградной кислоты), затем точно так же передается по цепочке белков-переносчиков и, наконец, торжественно "вручается" конечному получателю. Только в случае дыхания это опять-таки не хлорофилл, а другая молекула. Какая — попробуйте угадать. Ну конечно, это молекула кислорода! Получив необходимое количество электронов, ядовитый кислород тотчас присоединяет к себе соответствующее количество протонов (Н+) и превращается в безобидную воду. Так хитрая клетка убивает сразу двух зайцев: обезвреживает опасный яд и запасается энергией. Ведь система синтеза АТФ за счет энергии передаваемого "с рук на руки" электрона была унаследована дыхательной "молекулярной машинкой" от аппарата фотосинтеза (наш рассказ о появлении дыхания, конечно, весьма упрощен. В частности, считается, что эффективно использовать дыхание для получения энергии клетка научилась не в один миг. Поначалу, возможно, приходилось сжигать ценную органику "почти даром", только для того, чтобы обезвредить кислород). У цианобактерий до сих пор перенос электрона и в процессе фотосинтеза, и при дыхании осуществляется одними и теми же белковыми комплексами, так что между двумя процессами существует даже нечто вроде конкуренции за "право пользования" белками — переносчиками электронов. Однако появление свободного кислорода самый большой переполох произвело в третьем, нижнем слое сообщества. Местное население не умело фотосинтезировать, у них не было фотосинтетической электронно-транспортной цепи, чтобы быстро защититься от нового яда. Кроме того, метаболизм многих обитателей нижнего слоя было очень трудно приспособить к кислородным условиям. Например, для сульфатредукции и метаногенеза необходимы ферменты-гидрогеназы, которые в присутствии кислорода работать не могут. Бродильщикам было чуть легче. В конце концов они кое-как научились защищаться от небольших концентраций свободного кислорода при помощи специальных ферментов. Надо полагать, переход цианобактерий к оксигенному фотосинтезу происходил не в один миг, и концентрация кислорода в бактериальном мате росла более или менее постепенно. По мере совершенствования систем дыхания в верхних слоях сообщества кислородная угроза для нижних слоев снижалась (кислород "сжигался" при дыхании). В конце концов "наверху" появились аэробные гетеротрофы с таким эффективным дыханием, что кислород вообще перестал поступать в нижний слой. После этого жизнь наладилась. Так появились бактериальные маты "современного" типа. Нижний слой современных матов — бескислородный, и сульфатредукторы с метаногенами чувствуют там себя превосходно. Но в течение "переходного периода" анаэробным бактериям и археям нижнего слоя приходилось несладко. В качестве экстренной меры они стали активно заимствовать гены у других микробов (в последующих главах мы узнаем, что многие организмы целенаправленно пытаются хоть как-нибудь изменить свой геном в смертельно опасных ситуациях, и это служит мощным двигателем эволюции). В одной из групп архей массированное заимствование чужих генов пошло особенно удачно и приняло необыкновенно широкие масштабы. В конце концов это привело к появлению химерного организма: "сердцевина" у него осталась архейной, а почти вся "периферия" радикально изменилась и стала по большей части бактериальной. Изменился в том числе и обмен веществ. Наш химерный микроб стал, по-видимому, микроаэрофильным, то есть способным защититься от небольших концентраций кислорода и даже использовать кислород для некоторых биохимических реакций, но не способным дышать. Он умел сбраживать углеводы путем гликолиза, превращая их в пируват. Всему этому и многому другому он "научился" у бродильщиков, гены которых присваивал. Может быть, он даже научился хищничать, растворяя клеточные стенки других микробов и высасывая содержимое. Однако карьера специалиста по генетическому заимствованию требовала определенных изменений в механизмах работы с наследственной информацией. Самое интересное, что многие уникальные особенности эукариот могли развиться именно в связи с приспособлением к широкомасштабному присвоению чужих генов. Во-первых, множеством чужеродных генов нужно было эффективно управлять, отделяя "зерна от плевел", — это создало предпосылки для развития систем генной регуляции. Во-вторых, организм, столь "неразборчивый в связях", просто не мог не нахватать всевозможной генетической заразы — вирусов и мобильных генетических элементов (фрагментов ДНК, способных размножаться и перемещаться с места на место в пределах генома). О мобильных генетических элементах мы расскажем подробно в главе "Наследуются ли приобретенные признаки?". Мобильные элементы стали быстро размножаться в химерном геноме и прежде, чем клетка научилась обуздывать их активность, успели встроиться внутрь многих полезных генов. Так появились интроны — "бессмысленные" вставки в генах. Многочисленные интроны — одно из важнейших отличий эукариотической клетки (у прокариот они тоже встречаются, но их число на много порядков ниже). В-третьих, чтобы переполненные интронами гены продолжали нормально работать, нужна была эффективная система сплайсинга (от английского splicing — "сращивание, склейка встык") — вырезания интронов из "считанных" с генов молекул матричной РНК (мРНК). Это тоже важное отличие эукариот от прокариот. В-четвертых, сплайсинг — процесс медленный, а трансляция (синтез белка на основе мРНК) — быстрый. У прокариот трансляция начинается сразу после синтеза мРНК (транкрипции), часто даже до того, как этот синтез закончился. Рибосомы садятся прямо на недоделанную молекулу мРНК и начинают ее транслировать, пока РНК-полимеразы заканчивают транскрипцию. Если у вас в генах нет интронов, это нормально, но если интроны есть, нельзя допустить, чтобы они транслировались. Значит, нужно как-то отогнать рибосомы от недоделанных РНК, чтобы у последних была возможность спокойно завершить сплайсинг. Недавно было выдвинуто предположение, что именно это и было главным стимулом для формирования клеточного ядра. Ядерная оболочка отделила ядро ("область транскрипции и сплайсинга") от цитоплазмы ("области трансляции"). В-пятых, для нашей химеры было бы вполне разумно отказаться от хранения всего генома в форме единственной кольцевой хромосомы и перейти к системе из множества линейных хромосом. Так гораздо легче меняться генами со всеми подряд, перекраивать фрагменты генома, да и встраивать все новые и новые фрагменты ДНК в одно и то же разрастающееся кольцо вряд ли было бы удобно.
Линейные хромосомы — причина старения? Линейные хромосомы имеют один "недостаток" по сравнению с кольцевыми: при каждой репликации (копировании) они немножко укорачиваются, поскольку ДНК-полимераза не может скопировать самый кончик хромосомы, к которому она прикрепляется и с которого начинает репликацию. Кончики хромосом — их называют теломерами — не содержат полезных генов и состоят из коротких, многократно повторяющихся последовательностей нуклеотидов. С каждым клеточным делением теломеры укорачиваются. Когда теломеры становятся слишком короткими, клетка уже не может поделиться, не лишившись части полезной генетической информации. Поэтому клетки многоклеточного организма могут поделиться лишь ограниченное число раз. Возможно, в этом состоит одна из причин старения у многоклеточных эукариот (хотя эта гипотеза оспаривается многими экспертами). При образовании половых клеток теломеры приходится восстанавливать, достраивать. Для выполнения этой функции эукариоты приспособили "прирученные" мобильные элементы — ретротранспозоны (см. главу "Наследуются ли приобретенные признаки?"). Помимо всего перечисленного наша химера обладала еще одним уникальным свойством — она была способна к фагоцитозу, то есть умела "заглатывать" крупные объекты, в том числе целые бактериальные клетки. Современные бактерии и архей не умеют этого делать. Фагоцитоз сегодня характерен только для эукариот. Мы пока не знаем точно, когда и почему химерный предок эукариот приобрел эту способность. Может быть, он сначала стал фагоцитирующим хищником, и вместе с проглоченными микробами в его клетку стали постоянно попадать чужие гены, так что ему пришлось приспосабливаться к этому. А может быть, научился фагоцитировать, уже имея эффективные средства для работы с чужими генами. Так или иначе, в один прекрасный момент он проглотил маленькую пурпурную бактерию или, скорее, произошедшую от пурпурных бактерий гетеротрофную аэробную (умеющую дышать) бактерию и не стал ее переваривать. Этим знаменательным событием, собственно говоря, и завершилось в общих чертах становление эукариотической клетки. Наша полуфантастическая химера с живущими и размножающимися в ее цитоплазме аэробными бактериями — будущими митохондриями — это уже не что иное, как примитивная эукариотическая клетка. Митохондрии — органеллы, обеспечивающие энергией клетки всех животных и растений. Они размножаются делением, как бактерии, и не могут образовываться de novo, то есть "с нуля". Митохондрии унаследовали от своих предков-бактерий маленькую кольцевую хромосому, содержащую, правда, гораздо меньше генов, чем у любой бактерии. Геном митохондрии кодирует лишь малую часть белков, необходимых для жизни, нормального функционирования и размножения самой митохондрии. Все недостающие белки поступают в митохондрию извне — из цитоплазмы клетки, а кодирующие их гены находятся в клеточном ядре. На сегодняшний день твердо установлено, что практически все эти "митохондриальные гены ядерной локализации" когда- то располагались в митохондриальной хромосоме, а затем были перенесены в ядро. Быстрее всего процесс переноса шел на ранних этапах становления эукариотической клетки, то есть вскоре после того, как предки эукариот приобрели своих замечательных симбионтов. У животных этот процесс зашел дальше, чем у растений. Митохондриальная хромосома человека, например, кодирует всего 13 белков и имеет размер около 16,5 тысяч пар нуклеотидов. В ней остались только те гены, которые невозможно перенести по техническим причинам: строение кодируемых ими белков не позволяет транспортировать их через оболочку митохондрий. У растений митохондриальные геномы примерно в 10-20 раз больше, и эпизодический перенос отдельных митохондриальных генов в ядро продолжается по сей день. Возможно, еще до того, как произошло это эпохальное заглатывание, химера "всосала" в себя многие гены будущих митохондрий. Она ведь, разумеется, жила с ними бок о бок, в тесном симбиозе. Будущие митохондрии — обитатели второго слоя сообщества — защищали химеру, притаившуюся внизу, от избыточного кислорода. Химера производила низкомолекулярные углеводы (такие как пируват), а будущие митохондрии питались ими. Симбиоз мог зайти довольно далеко даже без объединения в единую клетку. Можно представить себе, что химера, перенявшая часть генов у своих аэробных соседей, постепенно начала снабжать их даже необходимыми белками. И постепенно, исподволь, научилась управлять их жизнедеятельностью — точно так же, как и сегодня эукариотическая клетка снабжает свои митохондрии белками и держит их под полным контролем. Симбионты, попав в такие "тепличные" условия, постепенно растеряли те гены, белковыми продуктами которых их обеспечивали извне. Активный перенос митохондриальных генов в ядро продолжался еще некоторое время после того, как химера "проглотила" предков митохондрий. Американские биологи недавно получили новые данные, показывающие, что этот процесс, вероятно, сильно замедлился после появления полового размножения и может вновь активизироваться в случае его вторичной утраты. Половое размножение препятствует переносу генов из митохондрий в ядро. V растений процесс переноса генов из митохондрий в ядро продолжается и по сей день. Остается открытым вопрос о том, зачем (или почему) митохондриальные гены переносятся в ядро. В долгосрочной перспективе ядерная локализация этих генов дает очевидные преимущества. К ядерным генам гораздо легче "пристроить" эффективные системы регуляции, которые позволяют увеличивать или уменьшать активность гена в зависимости от потребностей клетки. Ядерные хромосомы, в отличие от митохондриальных, у большинства организмов присутствуют в двух копиях (одна от отца, другая от матери). В процессе образования половых клеток парные хромосомы обмениваются между собой участками. В результате этого обмена (рекомбинации), а также вследствие слияния половых клеток (оплодотворения) в каждом поколении образуются новые сочетания генетических вариантов. Все это в конечном счете повышает генетический полиморфизм (разнообразие) популяции, ее устойчивость и приспособляемость к меняющимся условиям, создает более благоприятные условия для распространения полезных мутаций и снижает вероятность генетического вырождения вследствие необратимого накопления мутаций вредных. Митохондриальная хромосома не рекомбинирует. Митохондрии размножаются исключительно бесполым путем ("клонируются") и передаются только по материнской линии. Поэтому все потомки одной женской особи имеют одинаковые митохондриальные геномы, идентичные материнскому. Очевидно, это не очень "здоровый" способ передачи наследственной информации, практически исключающий возможность прогрессивной эволюции генов, оставшихся в митохондриях, при том что скорость накопления мутаций в митохондриальной хромосоме намного выше, чем в ядерных. Может быть, митохондриальные гены переселились в ядерный геном как раз для того, чтобы на них распространились все те преимущества, которые дают рекомбинация и половое размножение? Примерно так и рассуждают многие исследователи. Правда, в этом объяснении есть уязвимое место: преимущества, о которых идет речь, могут проявиться лишь в отдаленной эволюционной перспективе. Перенос гена из митохондрии в ядро едва ли может обеспечить организму или популяции мгновенную выгоду, а естественный отбор не в состоянии "заглядывать в будущее". Более того, этот перенос должен идти поэтапно, проходя целый ряд промежуточных стадий. Сначала копия митохондриального гена должна встроиться в одну из ядерных хромосом. Затем к ней должна в результате случайных перестановок участков ДНК пристроиться подходящая регуляторная область (чтобы ген заработал), а также особый фрагмент, который будет сигнализировать клетке, что белок — продукт данного гена — следует транспортировать в митохондрию. Все митохондриальные гены ядерной локализации имеют такой сигнальный фрагмент. Только после этого исходный ген, локализованный в митохондриальной хромосоме, может быть отключен или удален.
Многочисленные приспособления растений, препятствующие самоопылению, имеют, как выяснилось, еще одну функцию: они препятствуют крупным генетическим перестройкам. На всех этих промежуточных стадиях любые перетасовки генетического материала, происходящие в результате рекомбинации и полового размножения, могут только помешать делу. Например, представьте себе популяцию организмов, размножающихся половым путем, в которой у одних особей митохондриальный ген в ядерной хромосоме уже "включился", а у других еще нет, у одних митохондриальная копия еще работает, у других — уже нет. Ничего хорошего от скрещивания особей с разными состояниями этих признаков явно не получится, поскольку жизнеспособными будут не все, а только некоторые из возможных сочетаний ядерных и митохондриальных геномов. С гораздо большей вероятностью подобные генетические изменения возникнут и зафиксируются у организмов, размножаю щихся вегетативным путем или практикующих самооплодотворение. И это несмотря на то, что таким организмам перенос митохондриальных генов в ядро, казалось бы, вовсе не нужен, поскольку он не даст им тех долгосрочных преимуществ, о которых шла речь выше. Какой же фактор был важнее для переноса митохондриальных генов в ядро — долгосрочная выгода или сиюминутная возможность? Кстати, вопрос можно поставить и шире, ведь о многих эволюционных преобразованиях не так-то просто сказать, возникли они "зачем-то" или просто "почему-то". Биологи из Индианского университета в Блумингтоне (США) решили проверить эти гипотезы на растениях, у которых, как уже говорилось, перенос митохондриальных генов в ядро продолжается и по сей день. Если верна первая гипотеза, то есть гены переносятся ради долгосрочной выгоды, то у растений, практикующих перекрестное опыление, митохондриальные гены должны переноситься в ядро чаще, чем у самоопыляющихся или размножающихся вегетативно. Если же гены переносились не ради выгоды, а случайно, то это должно было происходить чаще у тех видов, которым это проще осуществить, то есть у самоопыляющихся или размножающихся бесполым путем. Ученые проанализировали геномы 170 родов покрытосеменных растений, время возникновения которых и положение на эволюционном древе более или менее твердо установлены. Для каждого рода было определено количество независимых событий переноса митохондриальных генов в ядро. Оказалось, что количество таких событий значительно выше в тех эволюционных линиях, где преобладают вегетативное размножение и самоопыление. Таким образом, подтвердилась вторая гипотеза: гены чаще переносятся не у тех видов, кому это выгодно, а у тех, кому это легче осуществить. Кроме того, оказалось, что корреляция между числом переносов и способом размножения лучше выражена для тех эволюционных событий (переносов), которые произошли сравнительно недавно, чем для более древних. Одна из возможных интерпретаций этого обстоятельства состоит в том, что отказ от нормального полового размножения — это своего рода эволюционный тупик, и растения, пошедшие по этому пути, либо довольно быстро вымирают, либо возвращаются к перекрестному опылению. Авторы предполагают, что тем же закономерностям должны подчиняться и перенос генов из пластид в ядро (пластиды, как мы помним, тоже являются потомками симбиотических бактерий), и перемещения генов с одной хромосомы на другую в пределах ядерного генома. Если эти предположения подтвердятся, это будет означать, что половое размножение является мощным фактором, препятствующим крупномасштабным геномным перестройкам у эукариотических организмов. (Источник: Yaniv Brandvain, Michael S. Barker, Michael J. Wade. Gene Co-Inheritance and Gene Transfer // Science. 2007. V. 315. P. 1685.) Конечно, описанный нами сценарий происхождения эукариот — лишь один из множества возможных, но некоторые его детали удивительно хорошо подтверждаются фактами. Например, удалось установить, что гены митохондриального происхождения на первых порах переносились в ядерный геном не по одной штучке в сто миллионов лет, а быстро и сразу большими порциями — в виде длинных фрагментов ДНК. То есть именно так, как, по нашим представлениям, должна была усваивать наследственный материал охочая до чужой ДНК химера. Последним актом в этой истории стало заглатывание химерой цианобактерии, которая дала начало пластидам. Наконец появившийся с такими трудами эукариотический организм мог вздохнуть с облегчением (ведь дышать-то он теперь умел — у него были митохондрии!). Он преодолел все трудности и стал независимым от микробного сообщества. С цианобактериальным фотосинтезом, альфапротеобактериальным дыханием, гликолизом "от бродильщиков" и эффективной системой управления геномом на архейной основе этот шедевр эволюционной "блочной сборки" теперь был сам себе полноценным сообществом. Впрочем, в первое время после своего появления эукариоты, возможно, оставались еще привязанными к своим родным бактериальным матам из-за недостатка кислорода в окружающей среде: цианобактерии еще не успели "надышать" его в достаточном количестве. Поначалу приличные концентрации кислорода можно было найти только в непосредственной близости от цианобактериальных матов. За пределами микробных сообществ кислород очень быстро вступал в реакции с различными растворенными в морской воде восстановленными веществами (особенно с двухвалентным железом). И пока все это железо не окислилось и не выпало в осадок, образовав крупнейшие залежи железных руд, концентрация кислорода в воде и воздухе оставалась низкой. Тут, конечно, пробил час железобактерий, о которых мы рассказывали в главе "Планета микробов", а также множества других хемоавтотрофных бактерий, быстро научившихся извлекать энергию из окисления самых разных веществ новым мощным окислителем — молекулярным кислородом. В ход пошли и соединения серы (серобактерии), и метан (аэробные метанотрофные бактерии). К сожалению, пока у нас слишком мало данных, чтобы построить точную хронологию описанных событий. Известно, что около 2,4-2,5 млрд лет назад происходил быстрый рост концентрации кислорода в гидросфере и атмосфере. Это называют "великим окислением" (great oxidation event). Впрочем, некоторые исследователи, в том числе известный палеонтолог А. Ю. Розанов, допускают значительно более раннее начало оксигенизации атмосферы (А.Ю. Розанов Ископаемые бактерии, седиментогенез и ранние стадии эволюции биосферы. 2003 http://evolbiol.ru/rozbakrus.htm ). 1,9-2,0 млрд лет назад, судя по молекулярно-филогенетическим реконструкциям и "молекулярным часам", жил последний общий предок всех современных митохондрий. Такой же возраст имеют и самые древние бесспорные ископаемые эукариоты. Однако есть и более древние находки, трактуемые некоторыми исследователями как остатки эукариотических клеток. Возможно, процесс становления эукариотической клетки растянулся на несколько сотен миллионов лет. Не исключено, что в течение этого переходного периода существовали многочисленные и разнообразные эукариотоподобные формы, в том числе с различными внутриклеточными симбионтами, но только 1,9-2,0 млрд лет назад возник особенно удачный симбиотический организм, потомки которого в дальнейшем вытеснили своих менее удачливых предшественников. Как бактерии превращаются в органеллыПосле приобретения митохондрий и пластид эукариоты вовсе не утратили способности к заглатыванию и "приручению" бактерий. Этот процесс продолжается и по сей день. Многие одноклеточные эукариоты (амебы, инфузории и другие) прямо-таки нашпигованы всевозможными симбиотическими прокариотами. Например, инфузории, обитающие в рубце жвачных, и жгутиконосцы, населяющие кишечник термитов, содержат в своей цитоплазме симбиотических бактерий, помогающих им переваривать клетчатку (целлюлозу). Такие симбиотические системы напоминают матрешку: в корове — инфузории, в инфузориях — бактерии. Зачем корова так тщательно пережевывает траву? Очень просто: кусочки травы должны стать достаточно маленькими, чтобы их могли проглотить симбиотические инфузории. Но и инфузория сама не может переварить целлюлозу и перепоручает это непростое дело своим собственным симбионтам — бактериям. Иногда бактерии и сами проявляют инициативу, изобретая различные способы проникновения в клетки эукариот. В этом случае велика вероятность того, что внутриклеточная бактерия станет не полезным симбионтом, а вредным паразитом (таковы, например, риккетсии). Впрочем, независимо от того, как будут складываться взаимоотношения внутриклеточных бактерий с хозяином, судьба их в некотором смысле предопределена. Жизнь внутри чужой клетки способствует постепенному упрощению и деградации. Внутриклеточные бактерии начинают идти по тому же пути, по которому когда-то прошли митохондрии и пластиды, то есть теряют свои гены, становятся все более зависимыми от хозяина и постепенно превращаются из самостоятельных организмов в нечто, очень напоминающее органеллы. У одних внутриклеточных бактерий этот процесс зашел уже очень далеко, у других он только начинается. Мы рассмотрим два самых крайних случая, в одном из которых внуктриклеточная бактерия, можно сказать, еще даже не начала упрощаться и терять гены, а в другом — практически уже стала органеллой, побив все рекорды генетической дегенерации. Первая из двух рекордсменок — бактерия Ruthia magnified, внутриклеточный симбионт, обитающий в тканях двустворчатого моллюска Calyptogena magnifica. Этот симбиотический "сверхорганизм" обитает на дне моря на большой глубине вблизи гидротермальных источников. Надо сказать, что дно океанов на глубине нескольких километров обычно довольно пустынно, и вовсе не потому, что живые организмы не выдерживают высокого давления. Лимитирующим фактором является пища: свет сюда не проникает, фотосинтез невозможен, и донные животные могут питаться лишь теми скудными крохами, которые падают сверху, из освещенных слоев воды, где жизнь гораздо богаче. Однако там, где из морского дна просачивается сероводород или метан, расцветают настоящие оазисы жизни. Существуют они за счет хемоавтотрофных бактерий, окисляющих H2S и СН4 при помощи кислорода (реже в качестве окислителя используются сульфаты или нитраты). Выделяющуюся в ходе этой химической реакции энергию бактерии используют для синтеза органических веществ из углекислого газа. Все животные, обитающие в гидротермальных оазисах (черви, моллюски, креветки), либо питаются этими бактериями, отфильтровывая их из воды, либо вступают с ними в симбиоз, поселяя бактерий на поверхности своего тела или даже внутри него. Сообщества морских гидротерм интересны тем, что они, в отличие от большинства других экосистем, существуют за счет энергии земных недр, а не солнечного света. Они почти независимы от окружающей биосферы, если не считать того, что кислород, образуемый где-то там наверху фотосинтезирующими организмами, им все-таки необходим.
Удачный симбиоз позволяет моллюску Calyptogena magnifica достигать весьма крупных размеров в условиях, где ни одно животное не способно выжить без помощи автотрофных бактерий. Автотрофные внутриклеточные симбионты гидротермальных животных по своей функциональной роли напоминают пластиды. Разница лишь в источнике энергии, используемой для синтеза органики: пластиды используют солнечный свет (то есть являются фотоавтотрофами), а гидротермальные бактерии — энергию химических реакций (хемоавтотрофы). Гигантский моллюск Calyptogena magnifica был одним из первых гидротермальных животных, описанных в научной литературе (его описали в 1980 году, вскоре после открытия глубоководных гидротерм). Пищеварительная система моллюска сильно редуцирована, и почти все необходимое он получает от живущего в его клетках бактериального симбионта. Симбиотическая бактерия Ruthia magnifica относится к группе гамма-протеобактерий (вместе со многими кишечными бактериями, включая кишечную палочку Escherichia coli, и разнообразными внутриклеточными симбионтами червей, насекомых и других животных). Геном бактерии был прочтен в 2007 году (L.G. Newton, T. Woyke, T.A. Auchtung et.al. The Calyptogena magnifica chemoautotrophic symbiont genome. // Science. 2007. v. 315. p. 998-1000). Как выяснилось, у Ruthia magnifica, несмотря на внутриклеточный образ жизни, генетическая дегенерация зашла совсем недалеко. Размер генома бактерии — 1,2 млн пар нуклеотидов. Для свободноживущих бактерий это маловато, но для внутриклеточных — очень много (больше, чем у всех прочих внутриклеточных бактерий с прочтенным геномом). У Ruthia magnifica, судя по набору генов, сохранились все метаболические пути, характерные для свободноживущих хемоавтотрофов. У нее присутствуют, например, все гены, необходимые для фиксации С02, для окисления восстановленных соединений серы, для синтеза различных кофакторов и витаминов, а также всех 20 аминокислот (в этом отношении Ruthia превосходит всех остальных изученных внутриклеточных микробов). Анализ генома подтвердил, что бактерия, как и предполагалось, фиксирует неорганический углерод при помощи цикла Кальвина (см. раздел "Самая главная химическая реакция" в главе "Планета микробов") — правда, с некоторыми "нетрадиционными" особенностями, — а энергию для этого получает за счет окисления соединений серы. Когда сероводород имеется в избытке, бактерия окисляет его до серы, которая в виде гранул хранится в бактериальных клетках "на черный день". Эти запасы впоследствии могут использоваться для дальнейшего окисления (до сульфита, а затем и до сульфата, который выводится из клетки специальными белками-транспортерами). В геноме бактерии имеются все необходимые гены для осуществления этих реакций. Моллюск-хозяин активно снабжает своего симбионта необходимой ему пищей: в крови моллюска обнаружен особый цинк-содержащий белок, предназначенный для связывания и транспортировки сероводорода. У бактерии есть также полный набор генов, необходимых для кислородного дыхания. Это означает, что микроб, как и предполагалось, использует в качестве окислителя кислород, которым его тоже обеспечивает хозяин. Имеются также наборы генов для таких важных метаболических путей, как гликолиз и цикл Кребса (Цикл Кребса — циклический биохимический процесс, происходящий в митохондриях эукариот, а также в цитоплазме некоторых протеобактерий. Является важным компонентом системы клеточного дыхания (наряду с электронно-транспортной цепью, о которой говорилось выше). Сначала цикл Кребса был не замкнут и служил для других целей; замыкание цикла произошло в связи с приспособлением протеобактерий к жизни в кислородных условиях). Все это есть и у хозяина, и бактерия могла бы просто брать готовые продукты извне, однако не делает этого. Бактерия снабжает своего хозяина аминокислотами и витаминами. Кроме того, она способна утилизировать отходы его жизнедеятельности, такие как аммоний. Это вещество бактерия всасывает из тканей хозяина и использует к обоюдной пользе, например, для синтеза тех же аминокислот. Главной особенностью Ruthia magnifica, конечно, является ее удивительная биохимическая самостоятельность. Такого полного набора генов, необходимых для основных биохимических процессов, свойственных свободноживущим хемоавтотрофам, ни у одной другой внутриклеточной бактерии не обнаружено. По-видимому, симбиотическая система Calyptogena magnifica — Ruthia magnifica находится на ранней стадии эволюционного становления и симбионт еще не успел далеко продвинуться по пути неизбежной в его положении деградации. Противоположный пример, показывающий, как далеко может зайти микроб по пути превращения в органеллу, дает бактерия Carsonella, о прочтении генома которой сообщила группа японских и американских исследователей в 2006 году (Atsushi Nakabachi, Atsushi Yamashita, Hidehiro Тoh, Hajime Ishikawa, Helen E. Dunbar, Nancy A. Moran, Masahira Hattori. The 160-Kilobase Genome of the Bacterial Endosymbiont Carsonella // Science. 2006. V 314. R 267.).
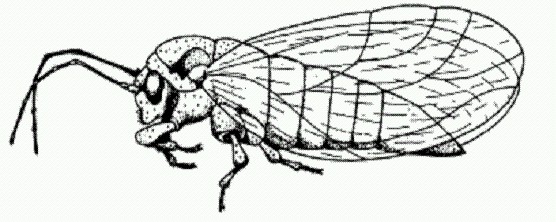
Листоблошки оказались обладателями удивительных симбионтов — бактерий с самым маленьким в мире геномом. Карсонелла живет в клетках листоблошек (Psyllidae) — мелких, похожих на тлей насекомых, питающихся исключительно соком растений. Как и другие насекомые, придерживающиеся этой более чем скромной диеты (например, тли и клопы), листоблошки обзавелись бактериальными помощниками, которые синтезируют для них необходимые вещества, отсутствующие в растительном соке, в первую очередь аминокислоты. Тут вернее будет сказать, что удачный симбиоз оказался решающим фактором, который позволил листоблошкам (и другим насекомым) перейти на питание чистым растительным соком. Как и Ruthia magnifica, карсонелла относится к группе гамма-протеобактерий. К той же группе относятся и многие другие внутриклеточные симбионты насекомых. Предполагается, что перед тем, как стать внутриклеточными симбионтами, предки карсонеллы жили в кишечнике насекомых. У карсонеллы наблюдаются все три основных признака генетической деградации, свойственной внутриклеточным бактериям: сокращение генома в результате потери почти всех некодирующих участков ДНК и значительной части генов, резкое преобладание в ДНК нуклеотидов А и Т и, соответственно, низкое содержание Г и Ц и быстрая молекулярная эволюция, то есть повышенный темп изменения ДНК в ряду поколений. По первому и второму пункту карсонелла побила все прежние рекорды. Ее геном втрое меньше, чем у архей Nanoarchaeum equitans, которая живет в гидротермальных источниках и паразитирует на другой архее — Ignicoccus (Это единственный случай, когда один прокариотический организм является облигатным (обязательным) паразитом другого)— и одной из разновидностей бактерии Buchnera, внутриклеточного симбионта тлей. У этих двух прокариот размер генома составляет 450-490 тысяч пар нуклеотидов (т.п.н.), и раньше именно они считались рекордсменами генетического упрощения. Карсонелла с ее 160 т.п.н. оставляет конкурентов далеко позади. Геном карсонеллы по размеру вполне сопоставим с геномом митохондрий. Типичные размеры митохондриальных геномов: 40-100 т.п.н. у низших эукариот, 200-400 т.п.н. у растений, 15-20 т.п.н. у животных. Само собой разумеется, что карсонелла не может жить вне клеток хозяина и передается только вертикально — от матери к ее детям (как и митохондрии). Сохранившихся у карсонеллы генов явно недостаточно для поддержания ее жизни — даже с учетом того, что она может пользоваться всеми благами внутриклеточного существования. Очевидно, что специализированные хозяйские клетки — бактериоциты — целенаправленно поддерживают жизнь симбионтов. Вполне возможно (хотя и не доказано наверняка), что многие гены, утраченные предками карсонеллы, были перенесены в геном хозяина, где они продолжают функционировать, обеспечивая бактерию необходимыми веществами извне. Как мы помним, именно это произошло когда-то с генами предков митохондрий. Вездесущий симбиозКогда-то симбиоз считался сравнительно редким явлением — скорее курьезом, чем правилом. Когда в 60-е годы XIX века было обнаружено, что лишайники представляют собой симбиотические комплексы из грибов и водорослей, ученые страшно удивились (надо же, какие причуды бывают у матушки-природы!). С тех пор многое изменилось. Уже в начале XX века отдельные выдающиеся мыслители предполагали, что симбиоз и кооперация могут играть огромную роль в развитии жизни на Земле. Хотя "организмоцентрический" подход в биологии по-прежнему господствует, сегодня ученые ясно понимают, что симбиоз — это магистральный путь эволюции, без которого прогрессивное развитие жизни на Земле было бы крайне затруднено, если вообще возможно. По-настоящему "автономный организм", сформировавшийся и живущий без всякого участия каких-либо симбионтов, в природе еще надо поискать. Большинство живых существ, населяющих планету, в действительности являются "сверхорганизмами" — сложными симбиотическими комплексами. Человек не исключение. Каждая наша клетка получает необходимую ей энергию от митохондрий — потомков симбиотических бактерий. Многие из наших генов получены нами от вирусов, всевозможных "эгоистических" фрагментов ДНК и мобильных генетических элементов (см. главу "Наследуются ли приобретенные признаки?"). Наш метаболизм во многом определяется многочисленными микробами, составляющими кишечную флору. И даже если заглянуть внутрь любого из этих микробов, то и там мы найдем сожителей-симбионтов (плазмиды, фаги, транспозоны). Кишечная микрофлора превращает человека в "сверхорганизм". В молекулярно-биологических исследованиях главный залог успеха — это удачный выбор объекта и методики. Новые эффективные методы, разработанные для решения конкретных задач, могут вдруг оказаться весьма полезными и в совершенно других областях. Именно это и произошло в данном случае. Американские биологи нашли необычное применение новейшим методикам, разработанным для "прочтения" геномов различных организмов. Ученые применили эти методы к ДНК, экстрагированной из человеческих фекалий, с целью получения общей характеристики кишечной микрофлоры. По имеющимся оценкам, в кишечнике взрослого человека присутствует более 1 кг микроорганизмов, относящихся к сотням различных видов. В точности их видовой состав неизвестен. Микробиологи знают "в лицо" лишь несколько десятков типичных представителей, которых можно вырастить на искусственных средах. Как выяснилось сравнительно недавно (и это открытие стало шоком для микробиологов), большинство существующих в природе микроорганизмов на искусственных средах не растет. Таких микробов называют некультивируемыми. В человеческом кишечнике они тоже, скорее всего, составляют большинство. Что-либо узнать об этих микробах удается лишь по нуклеотидным последовательностям ДНК в пробах, взятых из естественных сред. Исследователи выделили ДНК из фекалий двух человек, не принимавших целый год перед этим никаких лекарств, и провели масштабную работу по секвенированию — определению нуклеотидной последовательности фрагментов ДНК. Отсеквенированные кусочки затем собирались в более длинные фрагменты на основе наличия перекрывающихся участков. В итоге получилось около 74 тысяч неповторяющихся кусков общей длиной свыше 78 млн пар нуклеотидов. Для сравнения — в геноме человека свыше 3 млрд пар нуклеотидов, в геноме одной бактерии обычно 2-5 млн п.н. Разумеется, исследователи не рассчитывали получить полные геномы всех кишечных микробов. Для этого им пришлось бы приложить на несколько порядков больше усилий. Они лишь хотели получить общее представление о разнообразии микробного сообщества, его структуре и, главное, обмене веществ. А для этого полученной выборки геномных последовательностей оказалось вполне достаточно. Такой радикальный подход к изучению сообществ — свалить всех в одну кучу, истолочь и отсеквенировать — получил даже специальное название — метагеномный анализ.
Метагеномный анализ применяется в тех случаях, когда требуется выяснить, какие микроорганизмы присутствуют в пробе. Суть метода в тотальном выделении из образца (например, из морской воды, почвы, тканей какого-нибудь животного или из содержимого кишечника) всех молекул ДНК, какие попадутся. ДНК секвенируют (определяют последовательности нуклеотидов) и по этим последовательностям выясняют, какие существа присутствуют в пробе. Важное достоинство такого подхода в том, что он позволяет обнаружить и охарактеризовать микробов, не поддающихся культивированию в лаборатории (а таких среди микробов большинство). Генетические базы данных сегодня уже достаточно представительны, чтобы по набору выделенных из пробы генов можно было определить, какие организмы присутствуют в пробе, даже если эти организмы до сих пор не были известны науке. Сравнивая найденные гены с известными, можно выяснить не только чьими родственниками являются эти существа, но и как они живут и чем дышат. На следующем этапе в отсеквенированных последовательностях искали гены с известными функциями и пытались определить, какие фрагменты принадлежат бактериям, а какие — археям. Для этого последовательности сравнивались с известными, то есть внесенными в базы данных генами бактерий и архей, а также с их полными геномами. Отдельно анализировались гены рибосомной РНК (16S), по которым традиционно проводят классификацию микробов. В "явном виде" по генам рибосомной РНК удалось идентифицировать лишь 72 разновидности бактерий (из них 60 некультивируемых и 16 новых для науки) и один вид архей-метаногенов, однако авторы обосновали статистически, что, если бы работа по секвенированию ДНК из тех же самых проб была продолжена, число выявленных разновидностей микробов составило бы не менее 300. Оказалось, что в кишечной микрофлоре резко повышена доля генов, имеющих отношение к метаболизму полисахаридов растительного происхождения, некоторых аминокислот и витаминов, а также к метаногенезу. На основе проведенного анализа авторы выделили наиболее важные метаболические функции, которые выполняют микробы в человеческом кишечнике. Это прежде всего переваривание растительных полисахаридов, которые не могут перевариваться ферментами, закодированными в геноме человека. С этими трудноусваиваемыми углеводами расправляются в основном бактерии-бродильщики, выделяющие в качестве конечных продуктов обмена низкомолекулярные органические кислоты. Однако то, что для бактерий-бродильщиков является отходами жизнедеятельности, для человека — вполне съедобные вещества, которые активно всасываются кишечным эпителием. По имеющимся оценкам, из этого необычного источника люди получают около 10% калорий (эта оценка справедлива для приверженцев типичной "европейской" диеты). Задумайтесь, что это значит: не бактерии питаются нашими отходами, как кто-то мог бы подумать, а как раз наоборот! Кроме съедобных для человека веществ бактерии-бродильщики выделяют в качестве побочного продукта еще и молекулярный водород (Н2), который вреден для них самих и препятствует их росту и жизнедеятельности. Чтобы процесс переваривания растительных полисахаридов шел эффективно, кто-то должен постоянно утилизировать образующийся водород. Именно этим и занимаются археи-метаногены. В ходе метаногенеза поглощаются водород и углекислый газ и выделяется метан. В "совокупном геноме" кишечной флоры сильно повышено процентное содержание генов, связанных с синтезом незаменимых аминокислот и витаминов. Микробы сильно облегчают человеку жизнь, производя значительные количества этих необходимых нам веществ. Кроме того, кишечная флора располагает большим арсеналом ферментов для обезвреживания токсичных веществ, присутствующих в нашей повседневной пище, особенно растительной. Микробные геномы, таким образом, служат важным дополнением к геному Homo sapiens. Хоть это и нехарактерно для публикуемых в наши дни на Западе научных работ, авторы в данном случае решились на философское обобщение. По их мнению, человека следует рассматривать как "сверхорганизм", чей обмен веществ обеспечивается совместной слаженной работой ферментов, закодированных не только в геноме Homo sapiens, но и в геномах сотен видов симбиотических микробов. Между прочим, доля человеческих генов в совокупном геноме этого "сверхорганизма" составляет не более 1%. (Источник: Steven R. Gill et al. Metagenomic Analysis of the Human Distal GutMicrobiome // Science. 2006. V. 312. P. 1355-1359.) На симбиозе были основаны многие важнейшие ароморфозы (прогрессивные эволюционные преобразования). Мы уже познакомились с самым значительным из них — формированием эукариотической клетки, той основы, из которой в дальнейшем развились все высшие формы жизни (животные, растения, грибы и протисты (протисты - общее наименование всех одноклеточных эукариот)). В дальнейшем роль симбиоза в развитии жизни отнюдь не снижалась. Важнейшие функциональные блоки современной биосферы целиком и полностью держатся на симбиозе и симбиотических комплексах — "сверхорганизмах". Так, возможности высших растений — основных производителей органики и кислорода — были бы весьма ограничены без симбиоза с бактериями, способными переводить атмосферный азот в доступную для растений форму, и с почвенными грибами (микориза), без кооперации с насекомыми-опылителями и позвоночными — распространителями семян. Растительноядные животные — основные потребители производимой растениями органики — не могут эффективно переваривать растительную пищу без помощи разнообразных симбиотических бактерий и одноклеточных эукариот. Самые яркие и богатые жизнью морские экосистемы коралловых рифов невозможны без симбиоза коралловых полипов с одноклеточными водорослями — зооксантеллами. Сообщества различных экзотических, архаичных и экстремальных местообитаний (таких как наземные и подводные горячие источники, выходы метана и сероводорода, соленые лагуны, подземные воды и др.) тоже сплошь и рядом представляют собой сложные симбиотические комплексы микроорганизмов, в которых порой принимают участие и высшие организмы. Одними из самых важных являются так называемые азотфиксирующие симбиозы — кооперация растений с микроорганизмами, способными переводить азот из атмосферы или захороненной в почве органики в доступную для растений форму (аммоний, NH4+). Основная часть биосферного азота содержится в атмосфере в химически инертной молекулярной форме (N2). Восстановление (фиксация) этого азота требует огромного количества энергии. Как мы знаем из главы "Планета микробов", на это способны лишь некоторые бактерии и архей, у которых есть специальные ферменты — нитрогеназы. Дополнительная сложность состоит в том, что нитрогеназы работают только в анаэробных (бескислородных) условиях. Все высшие (эукариотические) организмы, в том числе растения по определению аэробны, и в этом, возможно, главная причина того, что у высших организмов способность к фиксации азота не встречается. Много азота содержится также в почве в составе органических веществ, но и этот азот для растений недоступен, поскольку у них нет пищеварительных ферментов, необходимых для расщепления этой органики. Два компонента азотфиксирующего симбиоза — это наземное растение (здесь годится любой тип растений) и какие-нибудь бактерии, способные фиксировать азот. В роли последних могут выступать цианобактерии, актинобактерии и альфапротеобактерии. Наиболее изучен симбиоз бобовых с клубеньковыми бактериями — ризобиями. Ризобии относятся к группе альфапротеобактерий. Ризобии, живущие в специализированных органах (клубеньках), снабжают растение аммонием, взамен получая весь комплекс элементов питания, в первую очередь углеводы, образуемые в ходе фотосинтеза. Между растительным и бактериальным компонентами симбиотического комплекса сложилась эффективная и гибкая система взаимной координации и регуляции. Например, специальные ферменты растений, работающие только в клубеньках, "заботятся" о том, чтобы концентрация кислорода в центральной части клубенька, где живут ризобии, была как можно ниже (и она там действительно ниже, чем в атмосфере, на 5-6 порядков). Биохимическая и генетическая интеграция симбиотического комплекса доходит даже до того, что активность некоторых растительных генов регулируется бактериальными белками! Важную экологическую роль играет также симбиоз растений с азотфиксирующими цианобактериями. В отличие от ризобий цианобактерии сами способны к фотосинтезу, что несколько упрощает задачу снабжения азотфиксирующих симбионтов необходимой энергией. Симбиотический комплекс водного папоротника Azolla и цианобактерии Anabaena имеет большое сельскохозяйственное значение: заселение рисовых плантаций этим папоротником резко повышает урожайность риса. Неслучайно в некоторых районах Юго-Восточной Азии азоллу обожествляют. В народнохозяйственных целях было бы очень полезно "научить" сами растительные клетки фиксировать атмосферный азот, точнее, их органеллы — пластиды. Теоретически это возможно, ведь многие "дикие" цианобактерии умеют фиксировать азот (мы ведь не забыли, что пластиды — потомки цианобактерий). Наверное, можно генно-инженерными методами создать пластиды с генами нитрогеназ, которые могли бы работать в темноте (например в корнях). Конечно, будет очень сложно добиться достаточно низкой концентрации кислорода в растительных клетках, но перспектива выглядит весьма заманчивой, ведь недостаток доступного азота — главный лимитирующий фактор, ограничивающий рост растений. Сняв это ограничение, можно было бы добиться колоссального увеличения урожайности.
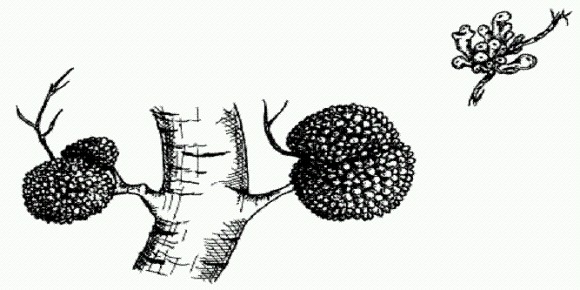
Клубеньки с азотфиксирующими бактериями встречаются не только у бобовых, но и у некоторых других растений. На рисунке — клубеньки на корнях ольхи. Огромную роль в биосфере играют также симбиозы автотрофов с гетеротрофами — кооперация организмов, синтезирующих органику из углекислого газа, с потребителями готовой органики. В роли первых выступают фотосинтезирующие организмы (растения, одноклеточные эукариоты, цианобактерии) или бактерии-хемоавтотрофы, использующие для фиксации СО2 энергию окисления неорганических веществ (например, сероводорода или метана). В роли вторых выступают животные или грибы. Широко распространены симбиозы с участием грибов — микоризы и лишайники. В случае микоризы грибной компонент получает от растения-хозяина углеводы (глюкозу, фруктозу), а сам берет на себя функцию корневых волосков (которые на микоризных корнях часто не развиваются) и вдобавок снабжает хозяина азотом и фосфором, которые гриб добывает, разлагая почвенную органику. Лишайники иногда называют "микоризой наоборот", поскольку в этих симбиотических комплексах гриб выступает в роли хозяина, а фотоситезирующие организмы (одноклеточные водоросли или цианобактерии) — в роли симбионта. Однако система биохимической интеграции у лишайников и микориз во многом сходна. Наибольшего совершенства эта система достигает у трехкомпонентных лишайников, в состав которых входят помимо гриба-хозяина специализирующиеся на фотосинтезе зеленые водоросли и специализирующиеся на азотфиксации цианобактерии.
Выход растений на сушу — результат симбиоза. Первые наземные растения появились в конце силурийского периода (более 400 млн лет назад). Недавно палеонтологи обнаружили, что уже самые первые наземные растения жили в симбиозе с грибами: у них была самая настоящая микориза. У этих растений еще не было настоящих корней — вместо них имелись так называемые ризоиды, не способные самостоятельно всасывать что-либо из почвы и служившие только для закрепления в грунте, а также, как теперь выяснилось, для обеспечения симбиоза с почвенными грибами. По- видимому, без этого симбиоза растения вообще не смогли бы выйти на сушу. Симбиоз растений с азотфиксирующими бактериями возник позже, причем для этого растениям оказалось достаточно лишь немного изменить те генетические системы, которые сложились у них ранее для взаимодействия с микоризными грибами. Симбиоз с автотрофами открывает большие возможности для многих водных животных, особенно малоподвижных (кишечнополостных, губок, асцидий, некоторых червей и моллюсков). Такие симбиотические комплексы представляют собой "сверхорганизмы", сочетающие признаки растений и животных (яркий пример — коралловые полипы). Автотрофы не только снабжают хозяина органикой, полученной в результате фото- или хемосинтеза, но и в ряде случаев помогают ему избавляться от конечных продуктов азотного обмена (например, мочевой кислоты или мочевины), которые служат для симбионтов ценным источником азота. Чрезвычайно широко распространены также симбиозы животных с микробами, помогающими усваивать растительную пищу. Потребление органики, производимой растениями в ходе фотосинтеза, — главная "экологическая роль" животных в биосфере, однако, как это ни парадоксально, сами по себе животные практически не способны справляться с этой задачей. Подавляющее большинство растительноядных животных попросту лишены ферментов для расщепления растительных полимеров (главным из которых является целлюлоза). Поэтому практически все животные-фитофаги — это на самом деле симбиотические комплексы из животного-хозяина и разнообразных бактерий, грибов или простейших (причем в последнем случае симбиотические простейшие зачастую сами имеют бактериальных симбионтов). Скорее всего, растительноядность изначально была симбиотическим феноменом. Роль симбионтов не сводится к расщеплению растительных полимеров: они могут также утилизировать азотные шлаки хозяина и синтезировать многие вещества, необходимые хозяину, но отсутствующие в растительной пище. Микробное сообщество, обитающее в пищеварительном тракте термитов, обладает даже способностью к азотфиксации, что позволяет этим насекомым питаться такими несъедобными вещами, как химически чистая целлюлоза. Некоторые биохимические процессы в таких симбиотических системах оказываются весьма причудливым образом распределены между хозяином и симбионтом. Например, комплекс "тли — бактерия Buchnera" синтезирует важнейшее вещество кофермент А совместными усилиями: сначала бактерия синтезирует из пирувата пантотеновую кислоту (чего не может насекомое), а затем тля синтезирует из пантотеновой кислоты кофермент А (чего не может бактерия). Конечным продуктом пользуются они вместе. Способность тлей приспосабливаться к колебаниям температуры зависит от симбиотических бактерий. Тли питаются исключительно соками растений. Жить на этой скудной диете им позволяет удачный симбиоз с бактериями Buchnera. Симбионты получают от хозяев кров и пропитание, а в обмен синтезируют для них аминокислоты, витамины и другие вещества, напрочь отсутствующие в той чуть сладенькой водичке, которая составляет единственную пищу тлей. То, что тли представляют собой в действительности симбиотические "сверхорганизмы", придает довольно своеобразный характер их экологии и эволюции. Например, у тлей вполне возможно наследование благоприобретенных признаков "по Ламарку". Ведь за жизнь одной тли в ее клетках сменяется несколько поколений симбионтов-бухнер, при этом генофонд бактерий может измениться в результате мутаций и отбора, и тогда тля передаст своим потомкам симбионтов, генетически отличных от тех, что она сама получила от матери (см. главу "Наследуются ли приобретенные признаки?"). Недавно сотрудники Факультета экологии и эволюционной биологии Аризонского университета в Тусоне (США) обнаружили, что генетические особенности симбионтов могут сильнейшим образом сказываться на жизнеспособности и экологических характеристиках всего симбиотического комплекса. Симбиозу тлей с бухнерами уже более 100 миллионов лет. За это время бухнера подверглась генетической деградации — это почти неизбежная судьба всех внутриклеточных симбионтов (конечным этапом деградации, как мы знаем, может стать превращение бактерии в клеточную органеллу). Бухнера потеряла способность к рекомбинации (обмену генами с другими бактериями), ее геном сильно сократился, упростился и потерял пластичность из-за утраты мобильных генетических элементов и повторов. Это снизило приспособительные способности бактерии. Как любой организм, привыкший к относительно неизменным благоприятным условиям, бухнера стала ужасно изнеженным и капризным существом. А это, в свою очередь, накладывает ограничения на весь "сверхорганизм". Например, бухнеры не любят жары — и это сильно мешает тлям распространяться в тех регионах, где высока вероятность перегрева. Между прочим, ограничения такого рода характерны и для многих других сверхорганизмов: мелкие наземные позвоночные не могли как следует освоить растительную диету, пока не выработали теплокровность (необходимая для переваривания клетчатки кишечная флора не выносит охлаждения); вспомним также, какие усилия прикладывают для поддержания постоянной температуры в своих жилищах термиты.
Компоненты симбиотического сверхорганизма: тля (справа) и бактерии Buchnera, живущие в специализированных клетках хозяина — бактериоцитах. Главная проблема, с которой организм сталкивается при перегреве, — это возможность денатурации (необратимого изменения структуры) некоторых белков. Обычно клетка в ответ на повышение температуры включает ряд генов (их называют генами теплового шока), кодирующих специальные белки — шапероны, чья функция состоит втом, что они "насильно" придают другим белкам правильную конфигурацию (см. сюжет "Когда вредных мутаций много, они не так вредны" в главе "Управляемые мутации"). Гены теплового шока (в количестве пяти штук) есть и у бухнеры. Любопытно, что некоторые гены, которые у других организмов обычно активируются при перегреве, у бухнеры всегда работают с повышенной активностью. Дело в том, что неправильное сворачивание белков может происходить не только из-за высоких температур, но и из-за вредных мутаций в генах, кодирующих эти белки. Бухнера, лишенная способности к рекомбинации, обречена на постоянное накопление мутационного груза, а шапероны до некоторой степени могут сгладить негативные эффекты мутаций, сворачивая правильным образом не совсем правильные белки. Ученые взяли три лабораторные линии гороховой тли Acyrthosiphon pisum и измерили активность генов бухнеры при нормальной температуре и при перегреве (чтобы определить активность генов, нужно выделить из клетки РНК и определить количество матричных РНК, считанных с каждого конкретного гена. Для этого существуют специальные приспособления - так называемые микрочипы). Оказалось, что у бухнер одной из трех линий при тепловом шоке не активируется ген ibpA, который кодирует небольшой "теплозащитный" белок, имеющийся не только у бактерий, но и практически у всех живых организмов. Поскольку линия тлей, где обнаружилась эта аномалия, была произведена всего пять лет назад от "нормальной" линии, в которой ibpA активируется при перегреве, ученые предположили, что причиной является мутация, возникшая менее пяти лет назад. Геном бухнеры прочтен, и найти мутацию было нетрудно. Исследователи отсеквенировали (определили нуклеотидную последовательность) регуляторной области гена ibpA и обнаружили, что у бактерий с аномальной реакцией на перегрев "потерялся" один нуклеотид (аденин) в том месте, где к хромосоме присоединяется регуляторный белок у32, который как раз и включает ген ibpA (и другие гены теплового шока) в стрессовой ситуации. В дальнейшем были проверены многие другие лабораторные линии тлей этого вида, и в одной из них обнаружился второй случай независимого возникновения той же самой мутации. Зная историю изучаемых линий, авторы установили, что один раз мутация произошла в 2001 году, второй — в 2005-м. Естественно, ученые захотели выяснить, встречается ли эта мутация в природных популяциях. Оказалось, что она довольно обычна у тлей из прохладных штатов Нью-Йорк и Висконсин (в некоторых популяциях мутантные симбионты присутствуют у 20% тлей), но не встречается в штатах Аризона и Юта, где бывает по-настоящему жарко. Эксперименты подтвердили, что у всех мутантов ген ibpA не активируется при перегреве, а у всех линий без мутации — активируется. Остальные гены теплового шока активировались совершенно одинаково как у мутантных, так и у нормальных бухнер. Исследователи решили выяснить, каким образом мутация, произошедшая у бактериального симбионта, отражается на жизнеспособности и репродуктивном потенциале симбиотического сверхорганизма. Тлей с мутантными и нормальными бухнерами выращивали в разных температурных условиях: при постоянной температуре 15 и 20°С, а часть насекомых в возрасте двух дней после вылупления из яйца подвергали четырехчасовому нагреванию до 35,5°С. Выяснилось, что после перегрева тли с мутантными симбионтами почти полностью утрачивают способность к размножению, тогда как тли с нормальными симбионтами размножаются вполне успешно. Однако при постоянной температуре 15 или 20°С заметным преимуществом обладают тли с мутантными симбионтами. Они раньше начинают размножаться и в среднем производят больше потомков. Дополнительные эксперименты позволили установить, почему кратковременный перегрев так губительно сказывается на тлях с мутантными бухнерами: повышение температуры до 35°С или выше приводит к массовой гибели бактериальных симбионтов. После этого остается всего лишь около 1000 бактерий на одно насекомое, тогда как норма составляет примерно миллион. Таким образом, мутантные бухнеры дают преимущество тлям при постоянной невысокой температуре среды, а "нормальные" выгодны в том случае, если существует опасность перегрева. По-видимому, в зависимости от климатических условий местности и даже от времени года направление отбора в природных популяциях тлей может меняться: иногда преимущество получают тли с обычными, а иногда — с мутантными симбионтами. В результате оба генетических варианта сохраняются в природных популяциях, и ни один из них не может полностью вытеснить другой. (Источник: Н. Е. Dunbar, А. С. С. Wilson, N. R. Ferguson, N. A. Moran. Aphid Thermal Tolerance Is Governed by a Point Mutation in Bacterial Symbionts // PLoS Biol. 2007. 5(5): e96.) Совместное существование позволяет каждому из компонентов симбиотического комплекса отказаться от тех биохимических функций, которые лучше удаются партнеру, и сосредоточиться на том, что лучше получается у него самого. Например, клубеньковые бактерии занимаются почти исключительно фиксацией азота, переложив заботу обо всем остальном на растение-хозяина. Часто один из партнеров специализируется на поставке в систему азота, а другой — углерода. По мере развития симбиотических отношений симбионт может полностью утратить самостоятельность и превратиться в неотъемлемую часть своего хозяина (впрочем, надо признать, что и хозяин при этом теряет самостоятельность и перестает быть "автономным организмом"). Грань между мутуалистическими (взаимовыгодными) и антагонистическими симбиозами — довольно условная и зыбкая. Например, многие растительно-грибные симбиозы в ходе эволюции могли долго "балансировать" на грани мутуализма и антагонизма, причем преобладающие потоки питательных веществ могли неоднократно менять свое направление. Непосредственный переход паразитизма в мутуализм — сранительно редкое явление (пример — "защитные симбиозы" растений со спорыньевыми грибами, в которых исходно паразитический гриб стал защищать растение от растительноядных животных путем синтеза токсичных веществ). В большинстве случаев симбиотические системы развиваются из фрагментов сообществ (экосистем). Например, симбиозы, основанные на биохимической кооперации, в большинстве случаев развиваются из кооперативных объединений свободноживущих организмов, соместно утилизирующих какой-то ресурс, или из фрагментов "трофической пирамиды" (симбиозы производителей органики с ее потребителями). Особый и весьма удивительный случай связи между трофической цепью и биохимическим симбиозом представляет морской моллюск Elysia viridis, питающийся водорослями. Этот моллюск ухитряется переселять пластиды съеденных водорослей в свои собственные клетки и долгое время сохранять их там живыми, приобретая таким образом способность к фотосинтезу. Настоящий гибрид животного и растения (В этом кратком обзоре важнейших типов симбиотических систем использованы данные из статьи Н. А. Проворова И Е. А. Долгих Метаболическая интеграция организмов в системах симбиоза // Журн. общей биологии. 2006. Том 67. № 6. Стр. 403-422. http://elementy.ru/genbio/resume/43 ; http:// elementy.ru/genbio/synopsis?artid=43.)! В трех заключительных разделах этой главы я расскажу о трех в высшей степени необычных симбиотических системах, которые ученым удалось недавно "расшифровать". Термостойкая траваФантастический случай тройного симбиоза описали в начале 2007 года американские биологи, работающие в Иеллоустонском национальном парке (США), где на горячей почве вблизи геотермальных источников произрастает термостойкая трава Dichanthelium lanuginosum, близкая родственница проса (Luis M. Marques, Regina S. Redman, Russell J. Rodriguez, Marilyn J. Roossinck A virus in a funfus in a plant: three-way symbiosis required for thermal tolerance. // Science. 2007. v.315. p. 513-515). Ранее было установлено, что удивительная устойчивость этого растения к высоким температурам каким-то образом связана с эндофитным (произрастающим в тканях растения) грибом Curvularia protuberata. Если выращивать растение и гриб по отдельности друг от друга, ни тот ни другой организм не выдерживает длительного нагревания свыше 38°С, однако вместе они прекрасно растут на почве с температурой 65°С. Кроме того, даже в отсутствие теплового стресса растение, зараженное грибом, растет быстрее и лучше переносит засухи. Продолжая исследование этой симбиотической системы, ученые обнаружили, что в ней есть еще и третий обязательный участник — РНК-содержащий вирус, обитающий в клетках гриба. Сначала ученые, конечно, не подозревали, что вирус важен для данного "сверхорганизма". Они просто решили выяснить, не оказывает ли обнаруженный вирус какого-нибудь влияния на взаимоотношения гриба и растения. Для этого они "вылечили" гриб, подвергнув его мицелий высушиванию и замораживанию при -80 °С. Эта суровая процедура приводит к разрушению вирусных частиц (и счастливы те организмы, которые могут, как грибы, сами ее выдержать и таким образом исцелиться от вирусных болезней!). Необходимые для экспериментов "безгрибные" растения получали из семян, с которых снимали оболочку, а затем полоскали 10-15 минут в хлорке. Выращенные из таких семян растения затем заражали (или не заражали) симбиотическим грибом, капая на них из пипетки взвесь грибных спор. Оказалось, что гриб, "вылеченный" от вируса, не в состоянии сделать растение термоустойчивым. Растения с таким грибом погибали на горячей почве точно так же, как и растения без гриба. Однако нужно было еще убедиться, что дело тут именно в вирусе, а не в каких-то побочных эффектах тех жестоких процедур, которые применялись при "лечении" гриба от вируса и растения — от гриба. Для этого "вылеченные" грибы были снова заражены вирусом, а этими повторно зараженными грибами, в свою очередь, заразили "вылеченные" растения. Теперь все было в порядке: заново собранный симбиотический комплекс отлично рос на горячей почве. Таким образом, для термоустойчивости оказались необходимы все три компонента симбиотической системы: и растение, и гриб, и вирус. Напоследок ученые провели совсем уж смелый эксперимент, свидетельствующий о высоком потенциале творческой энергии: взяли да заразили "грибом термоустойчивости" совершенно другое растение, а именно обыкновенный помидор. Были взяты четыре группы молодых томатов, по 19 растений в каждой. Первую группу заразили "дикой" формой гриба, содержащего вирус; вторую — грибом, вылеченным от вируса, а затем снова зараженным; третью — грибом, лишенным вируса; четвертую вообще оставили без грибов. Затем почву, в которой росли эти помидоры, стали каждые сутки нагревать до 65 °С на 10 часов, а остальные 14 часов температура почвы была 26°С. Спустя 14 дней в первой группе в живых осталось 11 растений, во второй — 10, в третьей — 4, в четвертой — только 2. Таким образом, гриб, зараженный вирусом, способен повышать термоустойчивость не только у своего природного хозяина — однодольного растения Dichanthelium lanuginosum, но и у неродственных растений, относящихся к классу двудольных. Это открытие может иметь большое практическое значение. Трудность пока в том, что авторам не удалось добиться стопроцентной зараженности всех помидоров симбиотическими грибами. Именно этим, по их мнению, объясняется более высокая смертность подопытных томатов на горячей почве по сравнению с Dichanthelium lanuginosum. Вместо выделительной системы — микробное сообществоЕще более фантастический случай симбиоза был описан в 2006 году исследователями из Германии и США (Woyke et.al. Symbiosis insights through metagenomic analysis of a microbial consortium. // Nature. 2006. 443. p.950-955). Объектом изучения стал малощетинковый кольчатый червь Olavius algarvensis, обитающий в Средиземном море. Червь этот интересен прежде всего тем, что у нет ни рта, ни кишечника, ни ануса, ни нефридиев — органов пищеварения и выделения. Некоторые другие морские черви тоже научились обходиться без органов пищеварения. Например, у погонофор кишечник превратился так называемую трофосому — тяж, набитый симбиотическими бактериями, окисляющими сероводород или метан. Поэтому можно было ожидать, что и у Olavius algarvensis отсутствие кишечника компенсируется наличием каких-то симбиотических микробов, обеспечивающих своего хозяина пищей в обмен на беззаботную жизнь в чужом теле. Однако редукция еще и выделительной системы — это явление беспрецедентное для кольчатых червей. Неужели микробы-симбионты сумели заменить червю не только органы пищеварения, но и органы выделения? Безусловно, этот случай заслуживал пристального изучения. Метагеномный анализ выявил присутствие в теле червя четырех видов симбиотических бактерий, два из которых относятся к группе гамма-протеобактерий, а два других — к дельта-протеобактериям. Обе гамма-протеобактерии, геном которых удалось ренконструировать почти полностью, являются автотрофами, то есть синтезируют органические вещества из углекислого газа. Необходимую для этого энергию они получают за счет окисления сульфида (S2-). В качестве окислителя используется кислород, а при отсутствии кислорода — нитраты (Использование нитратов в качестве окислителя для получения энергии называют нитратным дыханием. Нитратное дыхание широко распространено у бактерий, а недавно его обнаружили и у одноклеточных эукариот - фораминифер, родственников амеб). Если же нет под рукой и нитратов, окислителем могут служить некоторые органические вещества. В качестве конечных продуктов жизнедеятельности эти бактерии выделяют окисленные соединения серы (например, сульфаты).
Погонофоры и вестиментиферы ранее считались отдельным типом животных, но впоследствии было показано, что они являются сильно видоизмененной группой кольчатых червей. Сейчас их называют сибоглинидами. Они встречаются в больших количествах возле подводных гидротермальных источников, где много метана или сероводорода. Сибоглиниды являются по сути дела автотрофными животными. Если быть совсем точным, это автотрофные симбиотические "сверхорганизмы". Живущие в их трофосоме (бывшем кишечнике) хемоавтотрофные бактерии синтезируют органику из углекислого газа, а энергию для этого получают из химической реакции окисления сероводорода или метана. Червь питается органикой, производимой симбиотическими бактериями. Кровь червя переносит не только кислород, необходимый как червю, так и бактериям, но и сероводород — "пищу" бактерий. Жизнь погонофор почти совсем не зависит от остальной биосферы — все необходимое они получают из земных недр (за исключением кислорода, который производится оксигенными фотосинтетиками — растениями и цианобактериями). См.: В. В. Малахов. Вестиментиферы — автотрофные животные. 1997. http://www.pereplet.ru/obrazovanie/stsoros/394.html. Дельта-протеобактерии тоже оказались автотрофами, но другого рода, а именно сульфат-редукторами. Они получают энергию, восстанавливая сульфат (или другие окисленные соединения серы) до сульфида. Таким образом, метаболизм гамма- и дельта-протеобактериальных симбионтов оказался взаимодополнительным: отходы первых служат пищей вторым и наоборот. В качестве восстановителя (донора электронов, необходимого для восстановления сульфата) симбиотические дельта-протеобактерии могут использовать молекулярный водород. У них есть гены ферментов — гидрогеназ, необходимых для работы с молекулярным водородом. Возможно (хотя и не удалось доказать наверняка), что гамма-протеобактериальные симбионты производят некоторое количество Н2 и таким образом снабжают дельта-протеобактерий не только окисленными соединениями серы, но и восстановителем. Бактериальные симбионты живут не в глубине тела, а прямо под наружной оболочкой (кутикулой) червя. Здесь они ведут свою странную микробную жизнь, обмениваясь друг с другом продуктами своего метаболизма. Все прочее, чего им может недоставать, они получают из окружающей среды — в основном это вещества, просачивающиеся из морской воды под кутикулу хозяина. Микробы размножаются, а эпителиальные клетки червя тем временем потихоньку заглатывают их и переваривают. Этого источника питания, очевидно, червю вполне достаточно, чтобы не испытывать дискомфорта из-за отсутствия рта и кишечника. Но как удается червю обходиться без выделительной системы? Оказалось, что в геномах бактерий-симбионтов присутствуют гены белков, обеспечивающих всасывание и утилизацию мочевины, аммония и других отходов жизнедеятельности червя. Эти вещества служат бактериям ценными источниками азота. Возможно, основная выгода, которую бактерии получают от сожительства с червем, состоит в том, что он подвижен и может по мере надобности переползать туда, где условия среды наиболее благоприятны для всей честной компании. В верхних слоях осадка, где имеется немного кислорода, но нет сульфидов, гамма-протеобактерии могут получать необходимые им сульфиды от своих сожителей — дельта-протеобактерий. Сульфид в этом случае будет окисляться кислородом — наиболее энергетически выгодным окислителем. В больших количествах, правда, кислород вреден для сульфат-редукторов — дельта-протеобактерий. Если червь закопается поглубже, он попадет в слои, где кислорода нет вовсе. Здесь гамма-протеобактерии будут использовать в качестве окислителя нитраты, что несколько менее выгодно, зато сульфида у них будет вдоволь, потому что кислород больше не будет угнетать жизнедеятельность дельта-протеобактерий. Наконец, в еще более глубоких слоях осадка, где нет не только кислорода, но и нитратов, гамма-протеобактерии могут использовать в качестве окислителя некоторые органические вещества, выделяемые червем-хозяином и дельта-протеобактериями. При этом в клетках одного из двух видов гамма-протеобактерий запасается сера (как продукт неполного окисления сульфида), которую можно до- окислить позже, когда червь выползет повыше и станут доступны более сильные окислители. Щавелевая кислота, выделяемая гамма-протеобактериями, охотно утилизируется дельта-протеобактериями, и так далее: исследователи выявили еще целый ряд возможных способов "биохимического сотрудничества" внутри этого удивительного симбиотического комплекса. Таким образом, пять видов живых существ, объединившись, превратились в универсальный "сверхорганизм", способный жить в самых разнообразных условиях — в том числе и там, где ни один из его "компонентов" не выжил бы в одиночку. Лучшее — детям: как клопы кормят свое потомство ценными симбионтамиНу и напоследок совсем уж необыкновенный пример симбиотических отношений из жизни клопов. Об этом симбиотическом комплексе научный мир узнал в 2006 году благодаря исследованиям японских биологов (Takahiro Hosokawa, Yoshitomo Kikuchi, Naruo Nikoh, Masakazu Shimada, Takema Fukatsu. Strict Host-Symbiont Cospeciation and Reductive Genome Evolution in Insect Gut Bacteria // PLoS Biology. 2006. Volume 4. Issue 10.). Клопы семейства Plataspidae, или полушаровидные щитники, как выяснилось, не могут жить без симбиотических бактерий, обитающих в их кишечнике, — так же как и эти бактерии не мыслят себе жизни без клопов. Самка щитника, откладывая яйца, в каждую кладку помещает определенное количество "симбиотических капсул" — покрытых оболочкой шариков, содержащих бактерий-симбионтов в питательной среде. Вылупившиеся личинки первым делом поедают эти "пилюли", и бактерии попадают в задний отдел средней кишки. После этого происходят совсем странные вещи: средняя кишка пережимается посередине, так что передняя часть пищеварительной системы превращается в слепой мешок, не имеющий выхода. Пища перестает поступать в ту часть кишечника, где поселились бактерии. Питаются щитники соком растений, и все съеденное всасывается без остатка, а отходы жизнедеятельности затем выводятся из полости тела при помощи особых органов (мальпигиевых сосудов) прямо в заднюю кишку. Задняя часть средней кишки щитников превращается в инкубатор для бактерий. У самок этот отдел кишечника подразделяется на три части: в передней живут бактерии, в средней образуется питательный "наполнитель" для симбиотических капсул, а в задней образуется их оболочка. У самцов два последних отдела отсутствуют. Исследователи обнаружили, что без симбионтов щитники жить не могут. Удаление симбиотических капсул из кладки приводит к резкому увеличению смертности личинок и замедлению роста. У двух видов клопов (из четырех исследованных) все личинки погибли поголовно. У двух других видов часть личинок все-таки доросла до взрослой стадии, но получившиеся клопы отличались мелкими размерами, бледной окраской и были неспособны к спариванию. По-видимому, бактерии обеспечивают клопов необходимыми питательными веществами. Животные, питающиеся одним лишь соком растений, находятся в крайне трудном положении. Ведь в их пище практически отсутствуют жиры, белки, аминокислоты и многие другие необходимые вещества. На одних углеводах долго не протянешь. Правда, в растительном соке есть все элементы, необходимые для синтеза недостающих веществ, но справиться с такой сложной биохимической задачей способны только бактерии. Как мы уже знаем, другие насекомые, питающиеся растительными соками, такие как тли и листоблошки, тоже имеют бактерий- симбионтов, передающихся вертикально — от матери к детям. Главная особенность бактериального симбионта клопов-щитников состоит в том, что он живет не внутри, а вне клеток хозяина. Исследователи выделили из симбиотических капсул семи видов клопов молекулы ДНК бактерий-симбионтов и определили последовательность нуклеотидов в гене рибосомной РНК (16S рРНК). Этот ген традиционно используется для определения родственных связей бактерий. Оказалось, что симбионт клопов относится к группе гамма-протеобактерий, к подгруппе энтеробактерий, то есть кишечных бактерий (сюда же относится кишечная палочка), а ближайшим его родственником является та самая бухнера — симбионт тлей. Это само по себе весьма интересно, поскольку тли и клопы — родственные группы. Возможно, история клопино-бактериального симбиоза очень древняя и уходит корнями в те далекие времена, когда жили на свете общие предки тлей и клопов. На основе сравнения нуклеотидных последовательностей рРНК исследователи построили эволюционные деревья — отдельно для клопов и для их симбионтов. Эти деревья оказались абсолютно одинаковыми. Это означает, что эволюция клопов и их симбионтов протекала совершенно синхронно: появление нового вида клопа всегда сопровождалось появлением новой разновидности бактерии. Или, может быть, наоборот, изменение бактерии провоцировало появление нового вида клопа? Кроме того, это означает, что разные виды клопов не обмениваются между собой симбионтами. Последнее обстоятельство можно объяснить только физиологическими причинами, то есть тем, что каждая разновидность бактерии приспособлена только к клопам определенного вида, и наоборот. Дело в том, что разные виды полушаровидных щитников часто встречаются на одном и том же растении, и личинки вовсе не застрахованы от случайного поедания "чужих" капсул. Очевидно, такие ошибки плохо кончаются и для клопов, и для бактерий. Изученная японскими исследователями симбиотическая система представляет собой исключительно удобный объект, позволяющий без всяких усилий поставить множество интереснейших экспериментов. Например, что будет, если бактерий-симбионтов разных видов клопов поменять местами? Выяснить это проще простого — достаточно скормить личинкам чужие симбиотические капсулы. С внутриклеточными бактериями, такими как бухнера, провести подобный эксперимент крайне трудно или вообще невозможно. Наверняка этот и многие другие эксперименты будут поставлены в ближайшее время, и мы узнаем еще много увлекательных подробностей о жизни шестиногих симбиотических "сверхорганизмов". Что почитать на эту тему в ИнтернетеВ. А. КРАСИЛОВ. Нерешенные проблемы теории эволюции. 1986. http://evolbiol.ru/kr.htm В. А. КРАСИЛОВ. Метаэкология. 1997. http://evolbiol.ru/krasilov.htm (это насчет "эволюции с человеческим лицом") В. В. МАЛАХОВ. Вестиментиферы—автотрофные животные. 1997. http://www.pereplet.ru/obrazovanie/stsoros/394.html А. В. МАРКОВ. Обзор "Происхождение эукариот". http://evolbiol.ru/eucaryots.htm А. Ю. РОЗАНОВ, М. А. ФЕДОНКИН. Проблема первичного биотопа эвкариот. 1994. http://evolbiol.ru/ecorozanov.htm М. А. ФЕДОНКИН. 2003. Сужение геохимического базиса жизни и эвкариотизация биосферы: причинная связь. // Палеонтологический журнал. № 6. 2003. С. 33-40. http://evolbiol.ru/fedonkin2003.htm М. А. ФЕДОНКИН. Две летописи жизни: опыт сопоставления (палеобиология и геномика о ранних этапах эволюции биосферы). 2006. http://evolbiol.ru/fedonkin2006.htm |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Вверх |
||||
|
|
||||