|
||||
|
|
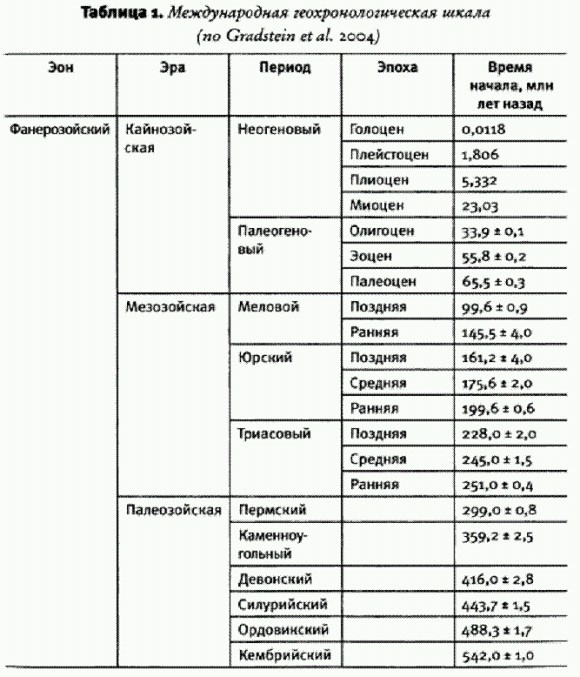
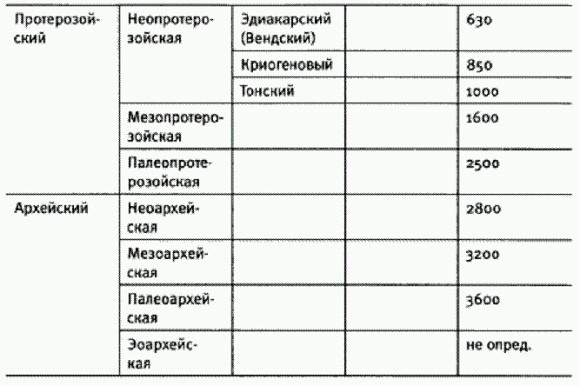
ГЛАВА 2. ПЛАНЕТА МИКРОБОВДревнейшие следы жизниВремя появления жизни на Земле точно не известно. Ясно одно: если наша планета когда-то и была безжизненной, то не очень долго. Земля сформировалась 4,5-4,6 млрд лет назад, но от первых 700-800 млн лет ее существования в земной коре осталось слишком мало следов. Главное, не сохранилось осадочных пород, в которых в принципе могли бы быть обнаружены следы жизни. Но имеются доказательства того, что гидросфера — водная оболочка нашей планеты — появилась очень рано. Об этом свидетельствуют, например, кристаллы циркона возрастом 4,4 млрд лет, обнаруженные в Западной Австралии. Строение и изотопный состав этих кристаллов позволяют предположить, что они сформировались в присутствии воды. Самым ранним свидетельством жизни считается облегченный изотопный состав углерода из графитовых включений в кристаллах апатита, найденных в Гренландии в отложениях возрастом 3,8 млрд лет. В этих включениях повышено процентное содержание легкого изотопа углерода 12С, что может быть результатом жизнедеятельности автотрофов — организмов, способных синтезировать органику из С02. Однако в ходе некоторых геологических процессов фракционирование изотопов углерода может проходить и без участия живых организмов. А это означает, что те древние кусочки графита, о которых идет речь, в принципе могли приобрести свой состав и в отсутствие доисторических существ.
Древнейшие ископаемые микроорганизмы возрастом около 3,5 млрд лет из Южной Африки внешне напоминают одноклеточных цианобактерий Synechococcus, хотя на внешнее сходство в данном случае едва ли стоит полагаться. Скорее всего, настоящие цианобактерии появились позже — 2,5-2,7 млрд. лет назад. Чуть более поздние отложения возрастом 3,7 млрд лет из той же Гренландии содержат в себе уже более достоверные следы жизни. Эти следы опять-таки представляют собой облегченный изотопный состав углерода, но в данном случае вероятность его абиогенного происхождения незначительна. Но какая это была жизнь — РНКовая или уже "современная", ДНК-РНК-белковая, доклеточная или клеточная, — определить невозможно. Однако можно уверенно сказать, что 3,55 млрд лет назад на Земле уже жили разнообразные микроорганизмы, напоминающие бактерий. В отложениях этого возраста появляются первые строматолиты — особые слоистые осадочные образования, формирующиеся в результате жизнедеятельности микробных сообществ. Здесь же найдены и сами окаменевшие микроорганизмы, напоминающие формой клеток некоторых современных бактерий. Это, конечно, ДНК-РНК-белковые клетки. С этого момента, собственно, и начинается палеонтологическая летопись как таковая. Самые древние (и самые интересные!) этапы становления жизни, включая эпоху РНК-мира, появление генетического кода и переход к ДНК-РНК-белковой жизни, к сожалению, не оставили внятных следов в земной коре. Поэтому их можно пока реконструировать только теоретически.
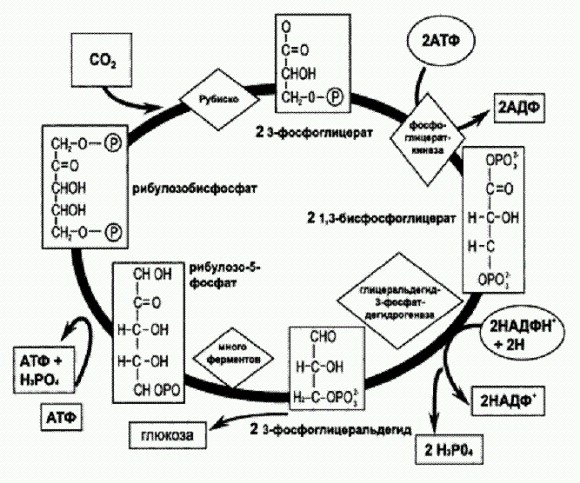
По форме клеток невозможно точно определить, к какой группе микробов относятся древнейшие ископаемые организмы, а кроме формы, от бактерий в палеонтологической летописи практически ничего не остается. Изредка, правда, удается найти "молекулярные окаменелости", или биомаркеры, — остатки некоторых органических молекул (более простых, чем ДНК, РНК и белки). Но и этого недостаточно для идентификации. Поэтому главным способом реконструкции древнейших этапов развития земной жизни сегодня является сравнительно-генетический анализ. Сравнивая между собой геномы современных микробов, ученые строят эволюционные "деревья", восстанавливая тот порядок, в котором происходило разделение эволюционных линий (для этого существуют весьма совершенные и сложные математические методики). Затем, зная примерную скорость накопления генетических изменений в разных участках генома, пытаются провести "калибровку" полученного древа, то есть датировать его узлы (точки разветвления). Палеонтологические данные тоже используются для калибровки там, где это возможно. Непрошеные помощникиРаздел написан в соавторстве с Еленой Наймарк. В течение очень долгого времени единственными живыми организмами на планете были прокариоты — бактерии и археи. Они встраивались в геохимические циклы, получая необходимую для жизни энергию за счет различных окислительно-восстановительных реакций. Последняя фраза, возможно, требует пояснений. Что значит "встраивались в геохимические циклы"? В поверхностных оболочках Земли — литосфере, атмосфере и гидросфере — как в древности, так и поныне происходит множество химических реакций и осуществляется круговорот веществ. Прокариоты с самого начала обладали уникальными высокоэффективными катализаторами — белками-ферментами, которые в принципе в состоянии катализировать (то есть многократно ускорять) чуть ли не любую мыслимую химическую реакцию. Если реакция идет с выделением энергии, эта энергия может быть "подхвачена" ферментами — АТФ-синтазами — и использована для синтеза АТФ. Имея запас АТФ, другие ферменты получают возможность осуществлять и такие химические реакции, которые идут не с выделением, а с поглощением энергии. В том числе синтез органики из углекислого газа. Вот, собственно, и весь секрет древней микробной жизни, ее химическая основа. Древнейшие прокариоты, скорее всего, были хемоавтотрофами (см. врезку). Они "пристраивались" к какой-нибудь химической реакции, которая шла и без их участия, сама по себе, только медленно. При помощи подходящего фермента они начинали катализировать эту реакцию, многократно ускоряя ее, а выделяющуюся энергию использовали для синтеза АТФ. Прокариоты и эукариоты, автотрофы и гетеротрофы. Биологи делят все живое (исключая вирусы, которых обычно не считают живыми) на три неравные части, называемые надцарствами: археи, бактерии и эукариоты. Первые две группы объединяют под общим названием "прокариоты". Прокариоты не имеют клеточного ядра, их геном находится прямо во внутренней среде клетки (цитоплазме) и обычно имеет вид единственной кольцевой молекулы ДНК (кольцевой хромосомы). У прокариот нет настоящего полового размножения, точнее говоря, в их жизненном цикле отсутствует фаза образования половых клеток и их попарного слияния в клетку с двойным набором хромосом — зиготу. У прокариот также нет внутриклеточных органелл, окруженных двойными мембранами, — митохондрий и пластид. Археи отличаются от бактерий в основном на молекулярном уровне. Внешне, по образу жизни или по способу получения энергии различить их довольно трудно. Правда, есть некоторые типы обмена веществ, характерные только для архей (например, метаногенез) или только для бактерий (например, кислородный фотосинтез). У архей по-другому устроены мембраны и клеточные стенки. У них, в отличие от бактерий, чаще встречаются интроны — некодирующие вставки в генах — и гистоны — специальные белки, участвующие в упаковке геномной ДНК. Архей чаще, чем бактерии, встречаются в различных экстремальных местах обитания. Например, есть архей, которые чувствуют себя комфортно в кипятке, а при 80°С начинают страдать от холода. Только среди архей встречаются микробы, паразитирующие на других микробах. Главные различия архей и бактерий — в нуклеотидных последовательностях их генов. Судя по величине этих различий, эволюционные линии бактерий и архей разделились чрезвычайно давно, на самой заре клеточной жизни. Эукариоты имеют клеточное ядро и окруженные двойной мембраной органеллы — митохондрии, служащие для кислородного дыхания, и пластиды, служащие для фотосинтеза (последние характерны только для растительных клеток). Доказано, что митохондрии и пластиды являются потомками симбиотических бактерий (см. главу "Великий симбиоз"). К эукариотам относятся разнообразные одноклеточные формы, обычно называемые "простейшими" (амебы, жгутиконосцы, инфузории, радиолярии и др.), а также многоклеточные — грибы, растения и животные. В жизненном цикле эукариот есть чередование гаплоидной и диплоидной фаз: пара гаплоидных (с одинарным набором хромосом) половых клеток сливается, образуя диплоидную (с двойным набором хромосом) клетку — зиготу. Это слияние двух половых клеток называют оплодотворением. Затем в какой-то момент происходит редукционное деление, или мейоз, в результате которого из диплоидной клетки образуются четыре гаплоидные. По способу получения органических веществ все организмы делятся на автотрофов и гетеротрофов. Организмы, умеющие превращать неорганический углерод в органические соединения, называются автотрофными,т.е. "самостоятельно питающимися". Организмы, не способные к этому, — их называют гетеротрофами — являются по сути дела нахлебниками автотрофов: они целиком и полностью зависят от производимых ими органических соединений. Автотрофы синтезируют органику из СО2, используя для этого энергию, полученную из какой-нибудь окислительно-восстановительной реакции (хемоавтотрофы) или путем фотосинтеза (фотоавтотрофы). Фотоавтотрофы, в свою очередь, делятся на аноксигенных (не выделяющих кислород) и оксигенных, или кислородных. Большинство архей — хемоавтотрофы, среди бактерий широко распространены все известные типы метаболизма, эукариоты являются либо оксигенными фотоавтотрофами (растения, одноклеточные водоросли), либо гетеротрофами (животные, грибы, многие простейшие). Подобные примитивнейшие экосистемы существуют и по сей день. Одну из них я имею удовольствие наблюдать каждое лето в северной Карелии, на берегу Белого моря, где провожу отпуск с семьей. Возле нашей избушки есть ручей, вытекающий из болота и бегущий к морю по дну глубокого оврага. Из крутых склонов оврага бьют ключи с ледяной прозрачной водой. Там, где ключевая вода смешивается с болотной, возникают и быстро растут комки мягкой скользкой рыжей мути довольно неприятного вида. Если долго нет дождей и течение в ручье замедляется, эта рыжая муть может заполнить все русло. Но достаточно хорошего ливня, чтобы вся эта гадость была смыта в море и ручей очистился. Если муть высушить, она превращается в кирпично-красный порошок, который отлично притягивается магнитом. По правде говоря, это самая обыкновенная ржавчина (Fe203), только склеенная чем-то слизистым и почти невесомым. Перед нами простейшая прокариотная экосистема. Мы видим естественный геохимический процесс, к которому "пристроился" автотрофный микроорганизм. Геохимический процесс в данном случае состоит в том, что подземные ключи, богатые недоокисленным растворенным железом (Fe2+), выходят на поверхность и соприкасаются с кислородом атмосферы. Кислород начинает окислять железо, которое превращается в нерастворимую ржавчину и выпадает в осадок. Этот процесс шел бы и без вмешательства микроорганизмов, только не очень быстро. Но за дело берутся непрошеные помощники — хемоавтотрофные железобактерии. Они многократно ускоряют процесс, а выделяемая ими "слизь" (она состоит в основном из углеводов) склеивает ржавчину в скользкие комки, которые нам и приходится долго разгонять, прежде чем набрать из ручья ведро воды. Впрочем, польза от этих бактерий тоже есть — именно они отвечают за образование так называемых "болотных руд", из которых можно даже выплавлять железо (если больше не из чего). В прошлом (особенно в протерозойскую эру) подобные бактерии играли ключевую роль в формировании крупнейших железорудных месторождений. Процесс до крайности неэффективен — чтобы увеличить свою биомассу на 1 грамм, бактерии должны окислить полкило железа. И цикл в данном случае не замкнут: недоокисленное железо поступает из земных недр, где его пока еще много, а окислившись, выпадает в осадок и сохраняется в таком виде неопределенно долгое время, накапливаясь в земной коре (в том числе в виде железных руд). И тем не менее система работает. Несмотря на всю ее примитивность, она обладает известным преимуществом — железобактерии практически не зависят от других живых существ. Они вполне самодостаточны, если не считать того, что используемый ими для окисления железа кислород производится не геохимическими процессами, а другими живыми организмами — оксигенными фотоавтотрофами (растениями, одноклеточными водорослями и цианобактериями). Одно из самых удивительных открытий в геологии за последние десятилетия состоит в том, что, как выяснилось, практически во всех геологических процессах, которые сформировали осадочный чехол нашей планеты, активно участвовали (и продолжают участвовать) микроорганизмы. Доказано, что многие месторождения руд — не только железных, но и золотых, марганцевых и многих других — имеют биологическое происхождение. Эти месторождения были некогда сконцентрированы микробами, постепенно осаждавшими на своих клеточных стенках ионы различных металлов. И если рассмотреть строение рудного вещества под микроскопом, становятся видны тельца, точь-в-точь такие, какими некогда были клетки микроорганизмов. В ходе своей жизнедеятельности микробы активно преобразуют соединения железа, серы, фосфора, образуя пириты, фосфориты и другие минералы. Как это происходит, не всегда понятно. Так что оценить масштабы этой четырехмиллиардолетней деятельности пока никто не берется. Между тем, зная механизм преобразования минералов микробами, можно было бы по внешнему виду минерала (в микромасштабе) и его составу отличить, создан ли минерал микроорганизмами или косной материей. Этот вопрос остро стоит, например, для марсианских минералов. Если бы удалось найти надежные признаки биологической активности в осадочных породах, то вопрос о жизни на Марсе был бы решен. Естественно, это касается и древней жизни на Земле. Но область эта еще очень слабо изучена. И начинать приходится с частностей. Месторождения цинка возникли благодаря бактериям. В осадочных породах иногда встречаются крошечные шарики сернистого цинка. Как выяснилось, в их образовании участвуют микроорганизмы. При извержении вулканов земная поверхность, будь то суша или морское дно, покрывается вулканическим пеплом. Этот пепел содержит очень много цинка, меди и свинца, которые затем не столько разносятся повсюду с водными потоками и выветриванием, сколько концентрируются в местах извержения. С течением времени вулканические лавы и пеплы при участии микроорганизмов могут превратиться в другие минералы, например в глины. Американские ученые из Калифорнийского университета и Национальной лаборатории имени Лоуренса в Беркли изучили материал из закрытой и заполненной водой шахты свинцово-цинкового месторождения Пикетт в юго-западном Висконсине. С 1999 года в тоннелях этой шахты вместе с микробиологами начали работу водолазы. Они увидели и засняли удивительную картину. В заброшенной шахте шла активная жизнь: стенки тоннеля были покрыты толстым слоем красно-оранжевой слизи и белыми сгустками. Это были различные анаэробные бактерии, получающие энергию с помощью окисления железа и преобразования серы. Оранжевый слой — цвет ржавчины — обозначал места деятельности бактерий, окисляющих железо, а белые сгустки — сульфатредукторов, восстанавливающих сульфат (SО42-) и использующих в своей жизнедеятельности ионы цинка. Наночастицы сернистого цинка — сфалерита — и окрасили эти пятна в белый цвет. Примерно такие бактерии работали на Земле в архейскую эру и продолжают работать сейчас в глубинах океанов, в глубоком почвенном слое, в рудных месторождениях. Ученые обнаружили, что в бактериальной биопленке образуются не только наночастицы сфалерита, но и относительно крупные (размером около микрона) шарики этого минерала. Каждый такой шарик состоит из множества наночастиц. Сформировавшись, шарик в силу своих солидных для молекулярного мира размеров становится менее уязвим для растворения и выноса и в результате остается в бактериальной пленке. Наночастицы имеют размеры в тысячи раз меньшие, сравнимые с размерами кластеров молекул воды. Поодиночке они были бы рассеяны в окружающем пространстве. Выяснилось, что цинковые шарики образуются при взаимодействии ионов цинка с белками и пептидами, богатыми аминокислотой цистеином (это одна из двух аминокислот, в состав которых входит сера). Ученые предполагают, опираясь на картину расположения клеток, органического вещества и цинковых сфероидов, что эти шарики организуются не за счет инкрустации стенок живых или мертвых клеток, а на скоплениях органической материи от распавшихся клеток. Цинковые наночастицы связываются с цистеином — таким образом образуется "затравка", к которой затем прикрепляются и цистеинсодержащие пептиды, и дополнительные цинковые наночастицы. В отсутствие цистеина крупные цинковые шарики не образуются. Все это ученые наблюдали под микроскопом, изучая и естественные руды, и материал, полученный в лабораторных опытах. Связывание цинка цистеином не было таким уж неожиданным для исследователей: ведь ферменты с высоким содержанием цистеина выполняют в клетке функцию поставщика необходимых ионов металлов — железа, марганца и др. Видно, в этот реестр входит и цинк. Теперь, обнаружив микрошарики сернистого цинка в осадочных породах, можно с полным основанием подозревать, что без участия бактерий тут не обошлось. Вот так в ходе, казалось бы, сугубо специальных исследований и познается прошлое и будущее жизни не только на Земле, но и на других планетах. (Источник: John W. Moreau, Peter К. Weber, Michael С. Martin, Benjamin Gilbert, Ian D. Hutcheon, Jillian F. Banfield. Extracellular Proteins Limit the Dispersal of Biogenic Nanoparticles // Science. 2007. V. 316. P. 1600-1603.) От самодостаточных эгоистов к единой биосфереПринято считать, что изначально в атмосфере Земли кислорода практически не было. Следовательно, железобактерии, с которыми мы познакомились в предыдущем разделе, не могли быть первыми, не с них началась на нашей планете "прокариотная эра". Судя по результатам сравнительно-генетических исследований, да и по логике вещей, первыми или, по крайней мере, одними из первых могли быть другие хемоавтотрофы — археи-метаногены. В простейшем случае они получают энергию, восстанавливая углекислый газ до метана при помощи молекулярного водорода: 4Н2+СО2 = СН4 + 2Н2О. Некоторое количество водорода образуется в земной коре в результате реакции перегретых горных пород с парами воды. Углекислый газ постоянно поступает из недр в атмосферу (например, при извержениях вулканов). Вот она, полная самодостаточность! Археям-метаногенам достаточно для жизни водорода, углекислого газа и воды (ну, конечно, в небольших количествах нужны еще азот, фосфор и всякие микроэлементы). Эти существа могли бы жить хоть на Марсе, хоть на полюсах Венеры, ведь они поразительно термостойки. Судя по сравнительно-генетическим данным, археи-метаногены появились около 3,8-4,1 млрд лет назад. Незамкнутость цикла в данном случае создает угрозу накопления метана в атмосфере, что может привести к сильному парниковому эффекту. Однако активность метаногенов на древней Земле, возможно, была ограничена. Лимитирующим фактором мог служить, например, недостаток молекулярного водорода. Самая главная химическая реакция. Жизнь на Земле основана на способности автотрофных организмов производить органику из углекислого газа (СО2). Поэтому тот химический процесс, в ходе которого СО2 включается в состав органических соединений, "фиксация СО2", по праву считается одним из важнейших (если не самым важным) в живой природе. Большинство организмов использует для фиксации СО2 циклическую последовательность химических реакций, известную под названием "цикл Кальвина". Ключевой фермент цикла Кальвина называется "рубиско" (это сокращенный вариант труднопроизносимого полного названия "рибулозобисфосфат карбоксилаза/ оксигеназа"). Рубиско — самый распространенный в мире фермент. Он присоединяет молекулу СО2 к органическому веществу, которое называется рибулозобисфосфат (сокращенно — RuBP). Таким образом молекула углекислого газа оказывается включенной в состав органических соединений. Недавно выяснилось, что у архей способ фиксации СО2 отличается от классического цикла Кальвина. Главное отличие состоит в том, что у архей в этом процессе непосредственное участие принимают рибонуклеотиды. Это хорошо согласуется с теорией РНК-мира. Фиксация СО2 — один из древнейших биохимических процессов, который должен был появиться еще в то время, когда все функции в живых организмах выполнялись молекулами РНК, рибонуклеотидами и их производными. Поэтому следовало ожидать, что и фиксация СО2 изначально должна была идти под контролем и при активном участии этих молекул. В цикле Кальвина участвует специальный фермент, ответственный за синтез RuBP. Этот фермент есть у растений и автотрофных бактерий, однако у архей он отсутствует. Поэтому до самого последнего времени было неясно, откуда архей берут RuBP, вещество, абсолютно необходимое для фиксации СО2. Эту загадку удалось разгадать японским биохимикам в 2007 году. Оказалось, что RuBP образуется у архей не из других фосфорилированных сахаров, как в цикле Кальвина, а из молекулы, являющейся составным блоком РНК и выполняющей "по совместительству" множество других функций в живой клетке, — аденозинмонофосфата (АМФ). АМФ — это рибонуклеотид, состоящий, как и все рибонуклеотиды, из азотистого основания (в данном случае аденина), сахара (рибозы) и одного остатка фосфорной кислоты. Есть все основания полагать, что этот способ фиксации СО2 эволюционно древнее, чем цикл Кальвина. Не случайно он обнаружен именно у архей — организмов, которых многие специалисты считают самыми архаичными формами жизни. (Источник: Takaaki Sato, Haruyuki Atomi, Tadayuki Imanaka. Archaeal Type III RuBisCOs Function in a Pathway for AMP Metabolism // Science. 2007. V. 315. P. 1003-1006.) В течение первого миллиарда лет существования прокариотной биосферы (примерно от 4,1 до 3,2 млрд лет назад) одни за другими появлялись новые формы микробов и новые способы получения энергии. В числе первых, по-видимому, появился бескислородный (аноксигенный) фотосинтез (? — 3,2—3,7 млрд лет назад). Его освоили бактерии — предки нынешних зеленых, пурпурных и цианобактерий. Аноксигенные фотосинтезирующие бактерии научились использовать энергию солнечного света при помощи особых светочувствительных молекул — бактериохлорофиллов и связанных с ними белковых комплексов. Молекулярная "машина", необходимая для осуществления эффективного аноксигенного фотосинтеза, устроена довольно сложно (хотя и намного проще, чем та, что используется оксигенными фотосинтетиками, появившимися позже). По-видимому, эффективный фотосинтез возник не сразу. Начиналось все с того, что некоторые микробы научились немного "подкармливаться" солнечным светом при недостатке других источников энергии. Для такой "подкормки" не нужны сложные молекулярные комплексы — достаточно одного-единственного светочувствительного белка. Недавно выяснилось, что подобные архаичные, простые и малоэффективные способы утилизации солнечного света до сих пор очень широко распространены в мире прокариот. Стало ясно, что способность к росту за счет энергии солнечного света (фототрофность) распространена в живой природе гораздо шире, чем считалось до сих пор.
Цикл Кальвина — важнейший биохимический процесс в живой природе, в ходе которого неорганический углерод вводится в состав органических молекул. При недостатке растворенной органики морские бактерии подкармливаются солнечным светом. Открытия последних лет показали, что кроме зеленых растений и цианобактерий, осуществляющих фотосинтез при помощи хлорофилла а, и известных ранее фототрофных бактерий, делающих то же самое при помощи бактериохлорофиллов, питаться солнечным светом могут и многие другие микробы, обладающие светочувствительными белками — протеородопсинами. Протеородопсины представляют собой своеобразные "насосы", локализованные в клеточной мембране и использующие энергию солнечного света для перекачки протонов (Н+) из клетки во внешнюю среду. Таким образом световая энергия переводится в разность электрохимических потенциалов, которая может затем использоваться клеткой для различных нужд, в том числе для синтеза АТФ. АТФ в свою очередь является универсальной энергетической "разменной монетой" клетки, используемой в самых разных энергоемких процессах. Протеородопсины были обнаружены у морских бактерий в 2000 году. Открытие привлекло большое внимание, поскольку оно означало, что многие организмы, до сих пор считавшиеся строго хемотрофными (получающими энергию за счет химических реакций), в действительности могут оказаться фототрофными — по крайней мере отчасти. В таком случае все представления об энергетическом балансе биосферы нуждаются в пересмотре. Фототрофность на основе протеородопсинов — это гораздо менее эффективный способ утилизации солнечной энергии по сравнению с настоящим фотосинтезом, но зато и гораздо более простой. Вскоре выяснилось, что протеородопсины встречаются не только у бактерий, но и у многих архей, обитающих в морях и океанах. По-видимому, морские микроорганизмы активно обмениваются друг с другом генами протеородопсинов, которые служат им важным подспорьем в конкурентной борьбе за энергетические ресурсы. Простота протеородопсиновой системы утилизации солнечного света облегчает ее горизонтальную передачу от одних микробов другим (о горизонтальном обмене генами подробно рассказано в главе "Наследуются ли приобретенные признаки?"). Однако до сих пор никому не удавалось экспериментально показать, что морские микробы, геном которых содержит гены протеородопсинов, действительно являются фототрофными организмами, то есть могут использовать энергию солнечного света для роста. Более того, первые попытки это подтвердить дали отрицательный результат: массовая и вездесущая морская бактерия Pelagibacter ubique, имеющая протеородопсины в клеточной мембране, в лабораторных условиях росла на свету не лучше, чем в темноте. Проблема осложнялась еще и тем, что большинство морских бактерий, содержащих протеородопсины, относятся к числу некультивируемых — в лабораторных условиях они просто не живут. Поэтому оставалось неопровергнутым предположение, что протеородопсиновая система использования солнечного света, возможно, настолько неэффективна, что не может внести заметного вклада в энергетический баланс морских экосистем и используется микробами лишь для каких-то второстепенных нужд. Лишь в 2007 году микробиологам из Швеции и Испании удалось показать, что это не так. Они обнаружили протеородопсины у бактерии Dokdonia, обитающей в Средиземном море. Бактерию удалось культивировать в лабораторных условиях. Оказалось, что в натуральной морской воде бактерия на свету растет гораздо лучше, чем в темноте. В первом случае плотность популяции после 100 часов инкубации составляла 3х105 клеток на миллилитр, во втором — в шесть раз меньше (0,5х105). Если культуру, выращенную в темноте, осветить, бактерии начинают быстро размножаться; если оставить в темноте — их численность постепенно снижается. Кроме того, бактерии, выросшие на свету, заметно крупнее тех, что росли в потемках. Дополнительные эксперименты показали, что влияние света на рост бактерий зависит от концентрации растворенной органики. Конечно, протеородопсины в отличие от систем настоящего фотосинтеза не могут сделать бактерию полностью автотрофной, то есть не нуждающейся в готовых органических веществах. Такие бактерии лишь "подкармливаются" светом и без готовой органики существовать не могут. Dokdonia практически не растет в воде с содержанием органики ниже определенного порога, и свет нисколько не помогает ей в этой ситуации. Однако при более высоких концентрациях растворенного органического вещества бактерия растет на свету в несколько раз быстрее, чем в темноте. Если же поместить докдонию в воду с избытком органики, то свет снова перестает влиять на ее рост (в этих условиях она растет одинаково хорошо как на свету, так и в темноте). Таким образом, протеородопсины повышают жизнеспособность морских бактерий лишь в условиях низких (но не слишком низких) концентраций растворенной органики. Возможно, отрицательные результаты, полученные ранее с Pelagibacter, объясняются именно нерациональным количеством органики в опытных средах. Так или иначе, мы теперь знаем, что многочисленные и разнообразные морские микробы, имеющие в своем геноме гены протеородопсинов, действительно могут быть факультативными фототрофами, то есть способны разнообразить свою диету, состоящую в основном из растворенной органики, солнечным светом. (Источники: 1. Laura Gomez-Consarnau et al. Light stimulates growth of proteorhodopsincontaining marine Flavobacteria // Nature. 2007. V. 445. P. 210-213; 2. Gazalah Sabehi et al. New insights into Metabolic Properties of Marine Bacteria Encoding Proteorhodopsins // PLoS Biol. 2005.3(8): е273.) Изобретение аноксигенного фотосинтеза было большим шагом вперед. Живые существа, овладевшие секретом фотосинтеза, получили доступ к неисчерпаемому источнику энергии — солнечному свету. Правда, их зависимость от дефицитных химических веществ, поступающих понемногу из земных недр, при этом все-таки сохранилась. Дело в том, что для фотосинтеза одного света мало — нужно еще какое-нибудь вещество, от которого можно оторвать электрон (это называется "фотоокисление"). В простейшем случае в роли донора электрона при фотосинтезе выступает сероводород. В результате деятельности аноксигенных фотосинтетиков сероводород превращается в серу (S) или сульфат (S042-). Опять незамкнутый цикл и накопление отходов! Но жизнь уже набирала силу, разнообразие микробов росло, и незамкнутые циклы постепенно начинали замыкаться. Планета захлебывается метаном и сульфатами? Что ж, эволюция нашла отличный выход из сложной ситуации: появились микроорганизмы, способные окислять метан при помощи сульфатов. Это были не просто микробы, а симбиотические микробные сообщества, состоящие из архей и бактерий. Архей окисляли метан, а бактерии восстанавливали сульфаты, причем оба процесса были каким-то не до конца еще понятным образом сопряжены между собой в неразрывное целое. Такие сообщества сохранились и по сей день в соответствующих местах обитания — там, где достаточно метана и сульфатов (например, в окрестностях подводных грязевых вулканов — см. ниже сюжет "В подводном грязевом вулкане обнаружены неизвестные микробы"). Результатом окисления метана был углекислый газ, необходимый всем автотрофам, а результатом восстановления сульфатов — сероводород, который с удовольствием использовали фотосинтетики. Циклы замыкались, биосфера приобретала устойчивость и способность к саморегуляции. Начиналась эпоха Великого Содружества Микробов. Примерно в тот же период (свыше 3,2 млрд лет назад), по- видимому, появились и первые гетеротрофы — так называемые бродильщики, которые получают энергию за счет бескислородной ферментации (сбраживания) готовой органики, произведенной автотрофами. В качестве отходов жизнедеятельности бродильщики имеют обыкновение выделять молекулярный водород, до которого в древней биосфере уже были охотники: во-первых, археи-метаногены, во-вторых, бактерии-сульфатредукторы (они охотно используют молекулярный водород в качестве восстановителя для восстановления сульфатов). Ясно, что на этом этапе большинство микроорганизмов уже не могли обходиться друг без друга. Даже в наши дни многие бродильщики наотрез отказываются расти в отсутствие микробов, утилизирующих выделяемый ими водород (сульфатредукторов или метаногенов), а тем, в свою очередь, жизнь не мила без бродилыциков. Уже 3,55 млрд лет назад на Земле, по-видимому, существовали сложные микробные сообщества — бактериальные маты. Именно они, скорее всего, ответственны за образование древнейших строматолитов. В наши дни нечто подобное можно наблюдать в некоторых экстремальных местообитаниях, таких как горячие источники. Древние бактериальные маты, вероятно, состояли из двух слоев. В верхнем обитали аноксигенные фототрофы. Они синтезировали органику из углекислого газа, потребляли сероводород и выделяли сульфаты. В нижнем слое жили бродильщики (они потребляли органику, произведенную фототрофами, и выделяли водород), сульфатредукторы (потребляли сульфаты и водород, производили сероводород), а также, возможно, метаногены с метанотрофами. В ходе жизнедеятельности сообщества под ним постепенно, слой за слоем, накапливался уплотненный осадок — так формировались слоистые образования, известные под названием строматолитов. Карбонат кальция — основной строительный материал строматолита — отчасти осаждался из морской воды, отчасти продуцировался самими микробами (в первую очередь сульфатредукторами). Реликтовые микробные сообществаВ реконструкции древнейших этапов развития микробной жизни большую роль играют исследования современных реликтовых микробных сообществ. Некоторые из них, как недавно выяснилось, могут существовать в полном отрыве от всей остальной биосферы в течение миллионов лет, получая все необходимое исключительно из земных недр. Одно из таких уникальных сообществ недавно было обнаружено глубоко под землей в Южной Африке. Все началось с того, что старатели на южноафриканском золотом прииске Мпоненг (Mponeng) стали бурить очередную скважину и на глубине 2,8 км наткнулись на водоносный слой. Глубинные воды, затерянные среди базальтов возрастом 2,7 млрд лет, находились под большим давлением, имели щелочную реакцию и оказались насыщены всевозможной химией: различными солями, среди которых преобладают сульфаты, растворенными газами, такими как водород, метан, углекислый газ и другие, и простыми органическими соединениями (углеводородами, формиатом, ацетатом). Большая часть органики, судя по изотопному составу, имеет абиогенное происхождение, то есть порождена не живыми организмами, а геологическими процессами. Температура подземной воды — чуть выше 60 градусов. Находка привлекла внимание микробиологов, изучающих биоту земных недр. На сегодняшний день хорошо известно, что толща земной коры заселена микроорганизмами вплоть до глубины в 6-7 км или даже более. Подземные микробы, по-видимому, играют большую роль во многих геохимических процессах, в том числе в образовании и деструкции нефти и газа. Неясным остается вопрос о том, в какой степени эта инфернальная микробиота является автономной, независимой от "внешней", большой биосферы, которая живет в основном за счет энергии солнечного света. Многие подземные микробы окисляют углеводороды или, к примеру, сульфиды при помощи кислорода, произведенного оксигенными фотосинтезирующими организмами (растениями и цианобактериями). Таких микробов, очевидно, нельзя назвать полностью автономными: исчезни жизнь на поверхности, и они тоже со временем погибнут. Другие — такие как археи-метаногены, восстанавливающие углекислый газ до метана при помощи водорода, — по-видимому, могли бы существовать в земных недрах неопределенно долго и после гибели всего живого на поверхности. Но до сих пор ни для одного подземного микробного сообщества не удавалось точно доказать, что оно в течение долгого времени действительно не использовало никаких веществ, произведенных "большой биосферой", и получало все необходимое исключительно из недр Земли. Микробиота, обнаруженная в воде из южноафриканской скважины, стала первым доказанным случаем долгого автономного существования живых организмов в недрах Земли без всякой связи с "большой биосферой". Об этом сообщила в октябре 2006 года группа исследователей из США, Канады, Германии, Тайваня и Южной Африки (Li-Hung Lin et al. Long-Term sustainability of a high-energy, low-diversity crustal biome // Science. 2006. v.314. p. 479-482.). Ученые обнаружили в подземных водах довольно большое количество микроорганизмов (40 млн клеток на литр). По нуклеотидным последовательностям генов рРНК, выделенных из проб, было установлено, что подавляющее большинство (более 88%) этих микробов относятся к одной разновидности сульфатредуцирующих бактерий. Эти бактерии получают энергию, восстанавливая сульфат (S042-) при помощи молекулярного водорода. Кроме сульфатредукторов в пробах обнаружены в небольших количествах и другие микробы — всего около 25 разновидностей, в том числе 4 разновидности архей-метаногенов. Расчеты показали, что в условиях, в которых живут эти микробы, сульфатредукция является наиболее выгодным из всех возможных типов энергетического метаболизма. Но микробиологический анализ был вовсе не главной частью исследования. Основное внимание авторы уделили изучению самой подземной воды, на которую была обрушена вся мощь современных методов аналитической химии. Воду разобрали чуть не по молекулам, изучили каждую примесь, измерили изотопный состав всех элементов, и все это для того, чтобы как можно точнее определить ее возраст и происхождение. Ученые пришли к выводу, что существует два возможных сценария образования этой воды. Согласно первому сценарию, вся она имеет поверхностное (атмосферное) происхождение и просочилась в недра в период между 15,8 ± 7,8 и 25 ± 3,8 млн лет назад. Вторая возможность заключается в том, что эта вода является смесью очень древней геотермальной воды возрастом 0,8-2,5 млрд лет и более "молодой" поверхностной воды, просочившейся на глубину 3-4 млн лет назад. В любом случае получается, что подземное микробное сообщество существует в абсолютной изоляции, без всякого притока вещества и энергии с поверхности, как минимум три миллиона лет, а возможно, и все 25 миллионов. И, несомненно, может существовать так и дальше — даже если все живое на поверхности погибнет, а из атмосферы исчезнет весь кислород. Изучение подобных реликтовых, изолированных от остальной биосферы микробных сообществ помогает понять, что представляла из себя жизнь на нашей планете миллиарды лет назад. Отважный странник. Когда эта книга уже готовилась к печати, стали известны новые удивительные подробности о жизни подземных микробов из прииска Мпоненг. Продолжая изучение подземной биосферы, исследователи выделили ДНК из 2600 литров воды, добытой в том же прииске и на той же глубине, но из другой скважины. К немалому удивлению ученых, в пробе обнаружился генетический материал лишь одного-единственного вида микробов. Анализы были проведены весьма аккуратно, и результат был подтвержден несколькими независимыми методами. Пришлось констатировать неожиданный и удивительный факт: обнаружена подземная экосистема, все население которой представлено лишь одним видом микробов. Следовательно, этот микроб должен обеспечивать себя всем необходимым без всякой помощи со стороны других организмов. Такая независимость и самодостаточность среди живых существ встречается очень редко. Что же это за микроб, живущий сам по себе в 60-градусной воде на глубине 2,8 км? Он оказался "старым знакомым", тем самым сульфатредуктором, который доминирует в пробах из прииска Мпоненг, изученных ранее. До сих пор, однако, об этом микробе было известно немногое (ему даже не было присвоено имя), и только теперь появилась возможность изучить его более основательно. Дело в том, что выращивать таких микробов в лаборатории практически невозможно, и судить об их биохимии, строении и образе жизни можно только на основе анализа ДНК. Однако если в пробе присутствуют геномы многих микроорганизмов, то понять, кому из них принадлежит тот или иной обрывок ДНК, технически очень сложно. Поэтому исследователи обычно ограничиваются анализом нескольких наиболее "показательных" генов, прежде всего — генов 16S-pPHK. По этим генам можно довольно точно определить, сколько и какие микробы присутствуют в пробе. Если попадается микроб, науке не известный, то по его гену 16S-pPHK можно определить, какой из известных микробов является его ближайшим родственником, а уже из этого делаются выводы о его биохимии и образе жизни. Другое дело, если в пробе присутствует только один микроб, — в этом случае современные технологии позволяют сравнительно быстро и недорого собрать из кусочков весь его геном. Именно это и проделали исследователи с уникальной бактерией. Имея в руках полную нуклеотидную последовательность генома, о микробе можно сказать очень многое. Для начала подземная бактерия получила имя — ее назвали Desulforudis audaxviator. "Audax viator" — слова из таинственной латинской фразы, указавшей герою повести Жюля Верна путь к центру Земли. В переводе они означают "отважный странник". Что ж, название вполне подходящее. По мнению исследователей, микроб совершил свое отважное путешествие в недра Земли и приспособился к жизни в полном одиночестве не менее 20 млн лет назад. Поскольку "отважный странник" в одиночку выполняет все функции, которые должны выполнять живые существа в экосистеме, авторы ожидали, что его геном должен содержать полный набор средств жизнеобеспечения в экстремальных условиях, включая биохимические механизмы для получения энергии, фиксации азота и углерода и синтеза всех необходимых веществ. Так и оказалось. В геноме D. audaxviator обнаружились следующие "рабочие инструменты": * полный набор генов для сульфатредукции, причем некоторые из этих генов бактерия явно позаимствовала когда-то у архей — микробов, по праву считающихся лучшими экспертами по выживанию в экстремальных условиях; * набор гидрогеназ — ферментов, позволяющих утилизировать молекулярный водород (который используется многими анаэробными микробами в качестве донора электрона в окислительно-восстановительных реакциях); * набор белков-транспортеров для перекачки готовых органических соединений — сахаров и аминокислот — из внешней среды в клетку. Это значит, что бактерия может вести себя не только как автотроф, то есть синтезировать органику из неорганических соединений, но и как гетеротроф, то есть питаться готовой органикой, если таковая вдруг появляется в окружающей среде. Очевидно, что там, где живет "отважный странник", единственным источником готовой органики могут быть мертвые распадающиеся клетки тех же самых бактерий. Не такова жизнь у "отважного странника", чтобы разбрасываться столь ценными ресурсами. Кстати, по имеющимся оценкам, бактерии, обитающие в подобных условиях, из-за острого дефицита ресурсов должны расти и размножаться невероятно медленно. Ученые не исключают, что между двумя клеточными делениями у таких микробов могут проходить сотни и даже тысячи лет; * белки для автотрофного метаболизма, позволяющие использовать в качестве источника углерода углекислый газ (СО2), угарный газ (СО) и формиат (НСОО-); * полный набор ферментов для синтеза всех 20 аминокислот; * гены, необходимые для формирования спор с плотной оболочкой (это, очевидно, позволяет "страннику" переживать периоды, когда условия становятся совсем уж невыносимыми); * гены, обеспечивающие образование жгутиков, при помощи которых микроб может плыть куда пожелает; * гены различных рецепторов и систем передачи сигналов, то есть того, что заменяет микробам органы чувств и нервную систему (по-видимому, "странник" чует, где выше концентрация дефицитных веществ, и плывет туда); * белки для транспорта аммония (NH4+) из внешней среды. В исследованных пробах концентрация аммония достаточно высока, чтобы полностью обеспечить микробов азотом, но, по всей видимости, так бывает не всегда. Поэтому D. audaxviator имеет в своем арсенале еще и нитрогеназу — фермент, позволяющий осуществлять азотфиксацию, то есть превращать молекулярный азот в удобоваримые для живой клетки азотистые соединения (прежде всего в тот же аммоний). Нитрогеназа, как и многие другие белки, была заимствована "странником" у архей путем горизонтального генетического обмена. Ученые нашли в геноме D. audaxviator немало других генов архейного происхождения. Большинство из них связано с приспособлением к жизни в экстремальных условиях. "Странник" позаимствовал у архей также и некоторые гены для защиты от вирусов (от них, оказывается, даже под землей не спрячешься). По-видимому, ему не удалось бы стать таким независимым и самодостаточным, если бы он предварительно не пообщался очень тесно с другими микробами и не одолжил бы у них кое-какие полезные гены. А вот чего у "странника" нет совсем, даже в рудиментарном виде, так это белков, позволяющих утилизировать кислород или хотя бы защищаться от его токсичного действия. Это значит, что с кислородом "страннику" не приходилось иметь дела уже очень давно. Главный теоретический вывод, сделанный авторами из изучения "отважного странника", состоит в том, что вся биологическая составляющая простой экосистемы, как выяснилось, может быть закодирована в одном-единственном геноме. (Источник: Chivian D. et al. Environmental Genomics Reveals a Single-Species Ecosystem Deep Within Earth // Science. 2008. V. 322. P. 275-278.) Первые альтруистыПо-видимому, уже на самых ранних этапах развития прокариотной биосферы микробам приходилось сотрудничать друг с другом, объединяться в сложные коллективы и сообща решать стоящие перед ними биохимические "задачи". Эффективность и устойчивость микробных сообществ повышались за счет развития средств коммуникации между микробами. Развивались системы химического "общения". Выделяя в окружающую среду различные вещества, микроорганизмы сообщали соседям о своем состоянии и влияли на их поведение. Тогда же зародился и альтруизм — способность жертвовать собственными интересами на благо сообщества. Возможно, многим читателям покажется сомнительным утверждение о существовании сложной социальной жизни, коммуникации и тем более альтруизма у микробов. Чтобы не быть голословным, приведу несколько фактов из жизни самой обычной, повсеместно встречающейся бактерии. Бактерия Bacillus subtilis — широко распространенный почвенный микроб, относящийся к числу наиболее изученных. Геном "тонкой бациллы" (так переводится с латыни название этого микроорганизма) прочтен еще в 1997 году, и функции большинства генов в общих чертах известны. Этого, однако, недостаточно, чтобы понять механизмы, управляющие сложным поведением бациллы. Этот микроб, например, умеет при необходимости отращивать жгутики и приобретать подвижность; собираться в "стаи", в которых передвижение микробов становится согласованным; принимать "решения" на основе химических сигналов, получаемых от сородичей. При этом используется особое "чувство кворума" — нечто вроде химического голосования, когда определенное критическое число поданных сородичами химических "голосов" меняет поведение бактерий. Мало того, В. subtilis способна собираться в многоклеточные агрегаты, по сложности своей структуры приближающиеся к многоклеточному организму. В критической ситуации (например, при длительном голодании) бациллы превращаются в споры, устойчивые к неблагоприятным воздействиям, чтобы дождаться лучших времен. Но превращение в спору для В. subtilis — процесс дорогостоящий, требующий активизации около 500 генов, и эта мера приберегается на самый крайний случай. Ну а в качестве предпоследней меры в голодные времена микроб прибегает к убийству своих сородичей и каннибализму. Если, конечно, сородичей вокруг достаточно много, то есть плотность популяции высока. Если нет, тогда делать нечего, приходится превращаться в споры натощак. Ученые выяснили, что при голодании у В. subtilis срабатывает особый генный переключатель, который может находиться лишь в одном из двух дискретных состояний (включено/выключено). "Переключатель" состоит из ключевого гена-регулятора Spo Активизация Spo До сих пор было неясно, почему токсин убивает только тех бацилл, которые его не выделяют (то есть тех, у кого Spo Таким образом, схватывание защитным белком молекулы токсина приводит к инактивации белка, тормозящего производство защитного белка. То есть чем больше будет токсина, тем больше клетка будет производить защитного белка. А как только токсин в окружающей среде закончится, молекулы SdpR перестанут инактивироваться, и производство защитного белка остановится. По молекулярно-биологическим меркам это крайне простой регуляторный каскад, проще некуда. Так бациллы защищаются от собственного токсина. А почему же бациллы с выключенным Spo Самым интересным тут является даже не каннибализм бацилл-убийц, а альтруизм бацилл-жертв, которые отключают себе все, что только можно, лишь бы помочь своим сородичам себя съесть. Казалось бы, естественный отбор должен способствовать закреплению в потомстве признака "Spo Сине-зеленые революционерыВажнейшим поворотным пунктом в развитии жизни стало изобретение оксигенного, или кислородного, фотосинтеза, благодаря которому в атмосфере начал накапливаться кислород и стало возможным существование высших организмов. Это великое событие произошло, по-видимому, 2,5-2,7 млрд лет назад (хотя ряд ученых придерживается мнения о более раннем появлении кислородных фотосинтетиков). "Изобретателями" кислородного фотосинтеза были цианобактерии, или, как их раньше называли, сине-зеленые водоросли. Как мы помним, при бескислородном фотосинтезе донором электрона служат соединения серы (чаще всего сероводород), а в качестве побочного продукта выделяется сера или сульфат. Недавно был открыт вариант бескислородного фотосинтеза, при котором донором электрона служат соединения железа. Побочным продуктом в этом случае являются более окисленные соединения железа. Не исключено, что именно микробы, осуществлявшие "фотоокисление" железа, ответственны за образование древнейших железных руд. Таким образом, существование аноксигенных фотосинтетиков зависит от довольно дефицитных веществ. Поэтому аноксигенный фотосинтез не мог обеспечить производство органики в количестве, необходимом для развития разнообразных гетеротрофов (потребителей органики), включая животных. При кислородном фотосинтезе донором электрона является обычная вода, а побочным продуктом — кислород. Изобретение кислородного фотосинтеза сделало бактерий независимыми от соединений серы или железа, и это открыло перед ними небывалые возможности. Ведь вода — ресурс практически неисчерпаемый. По сравнению с бескислородным фотосинтезом кислородный фотосинтез — гораздо более сложный процесс. Аноксигенные фототрофы утилизируют солнечный свет при помощи единого белкового комплекса, называемого фотосистемой. Для кислородного фотосинтеза потребовалось введение второго белкового светоулавливающего комплекса — второй фотосистемы. Обе фотосистемы в основных чертах похожи друг на друга (обе содержат хлорофилл, располагаются на клеточной мембране и отчасти состоят из похожих по структуре и функции белков). По-видимому, обе они являются вариациями одной и той же базовой "модели", то есть происходят от единого общего молекулярного "предка". Скорее всего, предки цианобактерий приобрели вторую фотосистему от каких-то других фотосинтезирующих микробов путем горизонтального переноса генов (см. главу "Наследуются ли приобретенные признаки?"). Объединившись в одной клетке, две фотосистемы со временем приспособились друг к другу, специализировались и разделили между собой функции. «Недостающее звено» в эволюции фотосинтеза. Как произошел переход от бескислородного фотосинтеза (при котором донором электрона служит сероводород) к кислородному, при котором донором электронов служит вода? Еще в 1970 году была предложена теоретическая модель, согласно которой этот переход осуществился через промежуточный этап, когда донорами электрона служили соединения азота (Olson J.M. The evolution of photosynthesis // Science. 1970. v.168. p. 438-446). Однако до самого последнего времени азотный фотосинтез оставался чисто гипотетической конструкцией — в природе его обнаружить не удавалось. Лишь в 2007 году азотный фотосинтез — искомый промежуточный этап на пути становления кислородного фотосинтеза — наконец-то был обнаружен. Открытие было сделано в ходе изучения микробов, обитающих в пресных водоемах и отстойниках сточных вод (Benjamin N. Griffin, Joachim Schott, Bernhard Schink Nitrite, an electron donor for anoxygenic photosynthesis // Science. 2007. v. 316. p. 1870). Микробиологи из университета Констанца (Германия) выращивали микробов в бескислородных условиях на свету в среде с небольшим количеством нитрита (N02-). Через несколько недель в 10 пробах из 14 стала заметна розовая окраска, характерная для бактерий, практикующих бескислородный фотосинтез, и было зарегистрировано окисление нитритов и превращение их в нитраты (N03-). При помощи специальных тестов удалось показать, что окисление нитритов является результатом именно фотосинтеза, а не какого-либо иного биологического или химического процесса. Активнее всего осуществляли "азотный" фотосинтез микробы, происходящие из отстойника города Констанца. Из этой культуры выделили самый массовый вид бактерий — шарообразные клетки диаметром 2-3 микрометра — и при помощи генетического анализа установили их родственные связи. Выяснилось, что ближайшим родственником этих микробов является Thiocapsa roseopersicina, широко распространенная фотосинтезирующая бактерия, относящаяся к группе пурпурных серных бактерий (эти микроорганизмы при фотосинтезе используют в качестве донора электрона соединения серы). Это открытие интересно еще и тем, что оно расширяет наши представления об участии микробов в круговороте азота. До сих пор не были известны фотосинтезирующие организмы, способные окислять соединения азота в отсутствие кислорода. Теперь эту возможность придется учитывать и при реконструкции ранних (бескислородных) этапов эволюции биосферы. Важность сделанного цианобактериями "открытия" трудно переоценить. Без цианобактерий не было бы и растений, ведь растительная клетка — результат симбиоза нефотосинтезирующего (гетеротрофного) одноклеточного организма с цианобактериями. Все растения осуществляют фотосинтез при помощи особых органелл — пластид, которые суть не что иное, как симбиотические цианобактерии. И неясно еще, кто главный в этом симбиозе. Некоторые биологи говорят, пользуясь метафорическим языком, что растения — всего лишь удобные "домики" для проживания цианобактерий. По сути дела цианобактерии не только изобрели кислородный фотосинтез, но и по сей день сохранили за собой "эксклюзивные права" на его осуществление. Цианобактерии не только создали биосферу "современного типа". Они и сегодня продолжают ее поддерживать, производя кислород и синтезируя органику из углекислого газа. Но этим не исчерпывается круг их обязанностей в глобальном биосферном круговороте. Цианобактерии — одни из немногих живых существ, способных фиксировать атмосферный азот (N2), переводя его в доступную для всего живого форму. Азотфиксация абсолютно необходима для существования земной жизни, а осуществлять ее умеют только прокариоты, и то далеко не все. Главная проблема, с которой сталкиваются азотфиксирующие цианобактерии, состоит в том, что ключевые ферменты азотфиксации — нитрогеназы — не могут работать в присутствии кислорода, который выделяется при фотосинтезе. Поэтому у азотфиксирующих цианобактерий выработалось разделение функций между клетками. Эти виды цианобактерий образуют нитевидные колонии, в которых одни клетки занимаются только фотосинтезом и не фиксируют азот, другие — покрытые плотной оболочкой гетероцисты — не фотосинтезируют и занимаются только фиксацией азота. Эти два типа клеток, естественно, обмениваются между собой производимой продукцией (органикой и соединениями азота). До недавнего времени ученые полагали, что совместить фотосинтез и азотфиксацию в одной и той же клетке невозможно. Однако новейшие исследования показали, что мы до сих пор сильно недооценивали метаболические способности цианобактерий. Эти микроорганизмы являются еще более универсальными и самодостаточными "биохимическими фабриками", чем было принято считать.
Цианобактерия Synecbococcus в процессе деления. Этот микроб днем фотосинтезирует, а ночью фиксирует атмосферный азот. В январе 2006 года Артур Гроссман и его коллеги из Института Карнеги (США) сообщили, что живущие в горячих источниках цианобактерии Synecbococcus ухитряются совмещать в своей единственной клетке фотосинтез и фиксацию азота, разделяя их во времени. Днем они фотосинтезируют, а ночью, когда в отсутствии света фотосинтез останавливается и концентрация кислорода в цианобактериальном мате резко падает, переключаются на азотфиксацию. Таким образом удалось выяснить, откуда берут азот микробные маты, живущие при температурах, не пригодных для роста обычных нитчатых азотфиксирующих цианобактерий с гетероцистами. Кроме того, открытие позволяет по-новому взглянуть на древнейшие этапы развития микробной жизни на нашей планете. После появления цианобактерий безраздельное господство прокариот на нашей планете продолжалось еще очень долго — полтора или два миллиарда лет. Микроорганизмы постепенно становились все более многочисленными и разнообразными (об этом можно судить по их ископаемым остаткам). Однако именно появление цианобактерий запустило цепочку событий, в результате которых эстафета эволюционного прогресса была в конечном счете передана более высокоорганизованным живым существам — эукариотам. Ключевую роль в этом сыграл кислород, накопившийся в атмосфере благодаря цианобактериям, а также давние традиции кооперации и симбиоза, сложившиеся в микробных сообществах еще на заре земной жизни. В конце концов уровень интеграции в сообществе прокариот достиг такого уровня, что несколько разных видов микроорганизмов слились в единый организм — эукариотическую клетку. О том, как это произошло, рассказывается в следующей главе. Что почитать на эту тему в ИнтернетеМ. В. ГУСЕВ, Л. А. МИНЕЕВА. Микробиология. 1992. http://evolution. powernet.ru/library/micro/index.html Г. А. ЗАВАРЗИН. Особенности эволюции прокариот //Эволюция и биоценотические кризисы. 1987. М.: Наука. С. 144-158. http://evolbiol. ru/zavarzin_1987.htm Г. А. ЗАВАРЗИН. Развитие микробных сообществ в истории Земли. 1993. // Проблемы доантропогенной эволюции биосферы. М.: Наука. С. 212-222. http://evolbiol.ru/zavarzin.htm Г. А. ЗАВАРЗИН. Эволюция микробных сообществ. 2003. (Доклад, прочитанный на теоретическом семинаре геологов и биологов "Происхождение живых систем". 15-20 августа 2003 г., Горный Алтай, стационар "Денисова Пещера"), http://evolbiol.ru/zavarzindok.htm А. В. МАРКОВ. Обзор "Зарождение жизни. Прокариотная биосфера 2003-2007. http://evolbiol.ru/paleobac.htm А. Ю. РОЗАНОВ. Ископаемые бактерии и новый взгляд на процессы осадкообразования. 1999. http://rogov.zwz.ru/Macroevolution/roza- nov1999.pdf A. Ю. РОЗАНОВ. Ископаемые бактерии, седиментогенез и ранние стадии эволюции биосферы. 2003. http://evolbiol.ru/rozbakrus.htm B. Н. СЕРГЕЕВ. Цианобактериальные сообщества на ранних этапах эволюции биосферы. 2003. http://evolbiol.ru/sergeev.htm М. А. ФЕДОНКИН. Две летописи жизни: опыт сопоставления (палеобиология и геномика о ранних этапах эволюции биосферы). 2006. http://evolbiol.ru/fed0nkin2006.htm
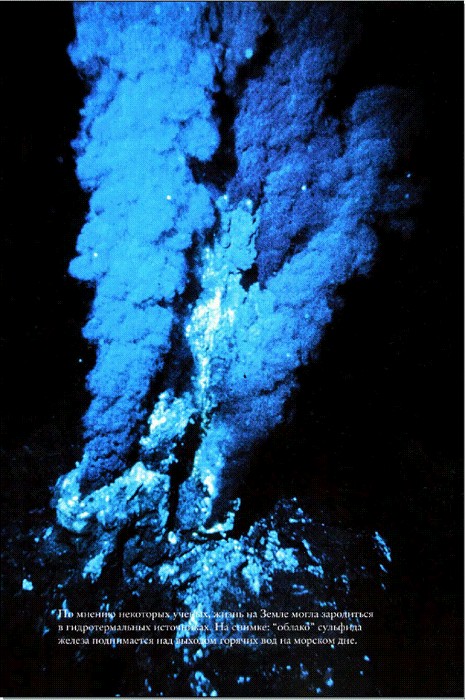
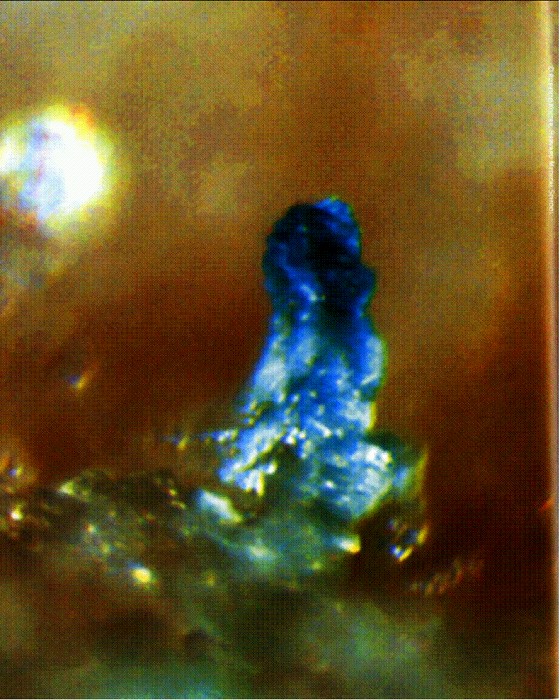
По мнению некоторых, ученых, жизнь на Земле могла зародиться в гидротермальных источниках. На снимке: "облако" сульфида железа поднимается над выходом горячих вод на морском дне.
Доисторический пейзаж.
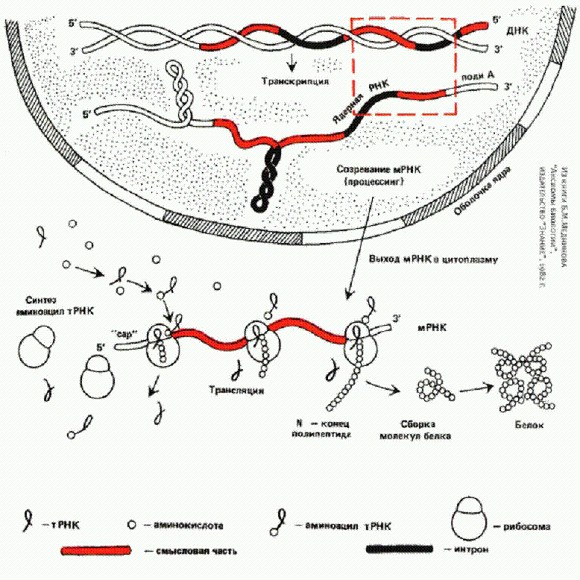
Схема обработки генетической информации в современной эукариотической (ядерной) клетке.
Рыжая муть на дне ручья и влажные комья ржавого цвета на берегу под хвощами — продукты жизнедеятельности железобактерий. Из этого материала в будущем может сформироваться болотная руда.
Прокариотный "оазис" на супралиторали (в зоне штормовых выбросов) Белого моря. Среди гниющих бурых водорослей на влажном соленом песке, насыщенном сероводородом, расцвели цианобактерии (зеленые пятна), пурпурные бактерии (розовые пятна) и белые серобактерии.
На этом снимке хорошо видны пузырьки фотосинтетического кислорода, образовавшиеся под слоем цианобактерий на беломорском песке. В центре — поселение пурпурных бактерий и раковинки мидий.
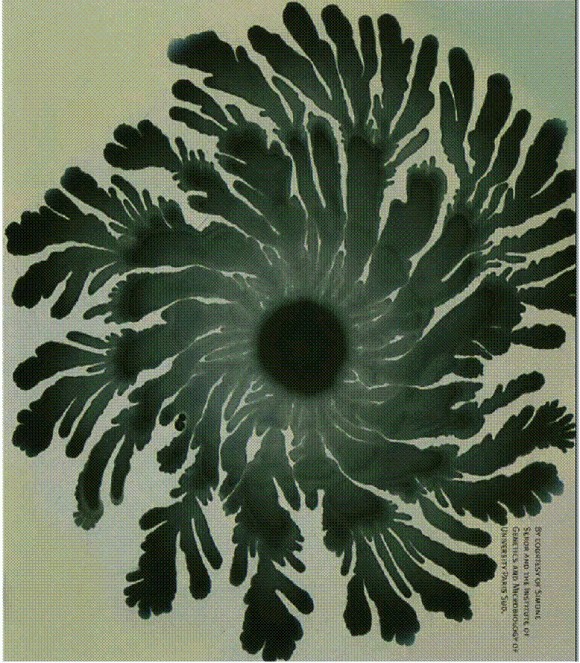
Многоклеточные агрегаты В. subtilis, образующиеся в результате сложного коллективного поведения, порой напоминают снежинки.
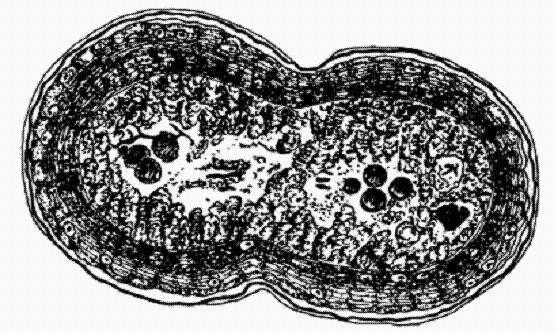
"Плодовое тело" Bacillus subtilis — сложный многоклеточный агрегат, часто образуемый дикими бактериями в природе и почти никогда — в лаборатории. |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Вверх |
||||
|
|
||||