|
||||
|
|
ГЛАВА ПЯТАЯРАННИЕ АЗИАТСКИЕ ВЕТВИ Все мы склонны подмечать различия. «Мамочка, почему этот дядя так не похож на того?» — подобные вопросы часто можно услышать из уст малышей. Итак, почему же люди, живущие на разных концах света, столь непохожи друг на друга? И почему взрослых и детей так занимает этот вопрос? Для многих и очень многих проблема этнических различий нашла свое выражение в идеях расизма, который является самым чудовищным и безобразным проявлением массового сознания. Но если попросту закрыть глаза на проблемы, окружающие нас со всех сторон, придется вечно топтаться на одном и том же месте. Этнические различия от этого вовсе не исчезнут. И лишь непредвзятые научные исследования, быть может, позволят нам выяснить, «почему этот дядя так похож на того» и почему нас тревожит вопрос о тех внешних различиях, которые характеризуют различные этнические группы. В предыдущей главе уже была в общих чертах изложена теория формирования этносов огромного азиатского континента. Она состоит в том, что многочисленные этнические группы, которые ныне заселяют просторы Азии, некогда распались на более мелкие, которые в свою очередь разошлись в разные концы обширного континента, что позволило им развиваться независимо друг от друга. Хотя распад крупного этноса на более мелкие кажется совершенно очевидным, а теория изолированного развития этнических групп получила научное подтверждение на основе данных генетики, однако нет пока достаточной ясности в том, каким был конкретно исторический процесс на азиатском континенте: ни окаменелости, ни археологические находки не раскрывают тайн прошлого. Дело в том, что археологические находки весьма немногочисленны, а окаменелостей сохранилось еще меньше, поэтому кажется преждевременным и спекулятивным любой ответ на вопрос, где предки различных современных этнических групп Азии обитали более чем за 70 тысяч лет до нас и когда они расселились на необозримых просторах этого континента. Окаменелости не позволяют получить точные ответы на поставленные вопросы, так обратимся же вновь к археологии, поскольку мы в состоянии датировать первобытные стоянки, опираясь на данные климатологии и генетики, а это в свою очередь поможет нам реконструировать маршруты передвижения первых жителей Азии. Пионеры проникали внутрь гигантского континента с трех сторон азиатского побережья Индийского и Тихого океанов. Выбор одного из трех направлений, изоляция и адаптация к новым условиям — все это сказалось на внешнем облике первопроходцев, глубоко врезалось в их черты и сделало их столь разными. Почему вопрос о внешних различиях волнует нас, как ни один другой? Что мы имеем в виду, когда говорим «разные»? Ответ на первый вопрос, возможно, коренится в самой природе человека, в том, что время отшлифовало в нас способность узнавать и помнить невероятное количество разных лиц. Нам не обойтись без этой способности отчасти уже потому, что круг нашего общения весьма широк[257]. Он значительно шире круга общения наших ближайших родственников из числа животных — шимпанзе, и взаимодействия между членами социальных групп у людей оказываются значительно сложнее. Мы должны уметь узнавать в лицо многих и многих людей. Оказавшись не в состоянии вспомнить имя того или иного человека, которое нам следовало бы помнить, мы смущаемся и краснеем, понимая, что допустили непростительную оплошность. Свою хорошую память на лица можно не только выгодно использовать, вращаясь в определенном обществе, но, обобщая и классифицируя свои наблюдения, можно выделить некие черты и внешние особенности, характерные для определенной группы людей, а затем найти различия между этой группой и какой-нибудь другой. Совершенно очевидно, что, разделив на основании внешних признаков людей на «своих» и «чужих», то есть тех, кто выглядит «по-другому», мы будем вести себя среди «своих» совершенно иначе, чем среди «чужих», относясь к последним с явным пренебрежением. К счастью, мы можем проникнуть внутренним взором в глубины своей совести и увидеть, что душа наша бессознательно стремится делить людей на «своих» и «чужих», что может привести к чудовищным преступлениям против человечности — к организованному расизму, и это понуждает пас следовать закону и усилием воли подавлять зародыши расизма в собственной душе и объявлять их вне закона. К сожалению, подобный самоконтроль не всегда увенчивается успехом. И мы оказываемся не в состоянии учиться на собственных ошибках, забывая злодейские погромы и травлю национальных меньшинств, забывая тех людей, что в прошлом стали жертвами расизма. Ограниченные возможности слова Побочным результатом борьбы с расизмом стала дискуссия о целесообразности признать табуированным само слово «раса», репутация которого непоправимо пострадала во времена нацизма. Поскольку в чистом виде рас не существует — они представлены многочисленными местными вариантами, переходными формами и смешанными разновидностями, многие антропологи считают понятие «раса» ненаучным, нарушающим права человека, бессмысленным и производящим впечатление некой абстракции, а следовательно, достойным признания вне закона. Все приведенные выше наблюдения весьма ценны, но факт остается фактом — и дети всегда его замечают: люди из разных регионов одного и того же континента могут иметь выраженные различия во внешнем облике. Короче говоря, ни запрет термина «раса», ни регулярная замена его различными эвфемизмами в такой ситуации не помогут. Наиболее часто в качестве альтернативы этому термину используются слова «население» или «этническая группа», но эти понятия столь расплывчаты, что могут лишь окончательно завести нас в тупик. Чтобы обозначить внешне отличающиеся типы людей, я использую в своей книге следующие термины — я вовсе не настаиваю на том, что они абсолютно точны, но зато они общеупотребительны и общепонятны, — итак, вот эти термины: «кавказцы» (или «кавказоиды»), «монголоиды» и «негритосы». Многие антропологи сочтут их неточными и отвергнут как явление, оскорбляющее личное достоинство человека. 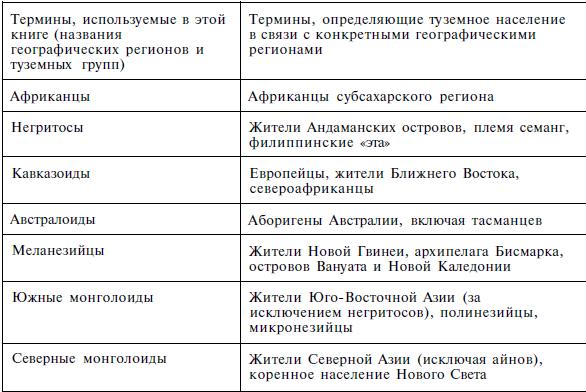 Рис. 5.0 Читатели, которые разделяют подобные взгляды, могут обратиться к таблице 5.0 — она позволяет перевести привычную для меня терминологию в систему терминов, основанную на географическом принципе и позволяющую обозначить туземное население, для которого характерен широкий спектр специфических особенностей строения организма через посредство зон его проживания. Хотя я и оставляю за собой право пользоваться привычной для меня терминологией, я вовсе не испытываю презрения к людям, чья внешность не похожа на мою собственную, — напротив, я прекрасно понимаю, что у всех прочих людей земного шара есть одно явное передо мной преимущество — ибо все они так не похожи на меня. Но вместе с тем я уверен, что политически корректные эвфемизмы и более современные термины могут скорее завести в тупик, нежели прояснить суть дела. Всем, конечно, известно слово «монголоиды» — оно часто используется для краткого обозначения целой категории людей, наделенных определенными чертами внешности и телосложения. Однако необходимо всегда помнить, что «традиционные» термины — это лишь неточные обозначения сложных жизненных явлений, которые не могут существовать вне бесчисленных конкретных вариаций в пределах определенной группы и вне системы взаимовлияний между различными народами. Самым очевидным дифференцирующим признаком, позволяющим отличать одни народы Евразии по внешнему виду от других, является цвет кожи, которая в более солнечных тропических регионах приобретает темный шоколадный оттенок. И это не случайное совпадение обстоятельств. Хотя гены еще таят в себе немало загадок, мы можем утверждать, что темный цвет кожи, обязанный своим появлением пигменту меланину, определяется генетической памятью человека, которая формировалась в процессе длительной эволюции. Кожа людей, которые живут в тропиках и субтропиках, подвергается мощной атаке ультрафиолетовых лучей. В результате этого человек рискует пострадать от чрезмерного загара и покрыться волдырями, а также умереть от рака кожи. Лишь темный цвет кожи способен уменьшить риск и угрозу для здоровья человека. Среди других, хотя и не столь очевидных преимуществ, можно назвать то, например, что пигментация, в которой участвует меланин, позволяет коже более эффективно противостоять перегреву и излучать излишки тепла, а также препятствует разрушению в организме такого важного витамина, как фолиевая кислота. Итак, в результате длившейся многие и многие поколения эволюции кожа людей в жарких климатических поясах стала темной. Именно темнокожие среди множества своих чад и домочадцев в областях повышенной солнечной активности живут дольше и чувствуют себя как нельзя более счастливыми[258]. В Северной Азии (т.е. в той части Азии, которая охватывает территории от Цзиньхай-Тибетского плато на севере и до Урала на востоке) и в Европе солнечный свет не столь интенсивен, и риск развития рака кожи менее выражен, зато население этих регионов повсеместно страдает от рахита и различных заболеваний костей, вызванных недостатком солнечного света. В самом Лондоне, причем уже в начале двадцатого столетия, от этих заболеваний умерло множество детей. Я сам был очевидцем подобных трагедий. Дело в том, что, попав как-то раз в больницу, а именно в отделение патологии Лондонского королевского госпиталя, я специально узнавал, насколько распространен рахит в наши дни. Во второй половине двадцатого столетия в Великобритании вновь получили широкое распространение такие заболевания, как рахит и остеомаляция[259], вторгшиеся в наши широты вместе с переселенцами с Индийского субконтинента. Именно дети потомственных индийцев чаще всего страдают рахитом. Это отчасти объясняется тем, что их темная кожа продолжает фильтровать солнечный свет, как это было необходимо на родине их предков, а не здесь, где солнечного света и без того недостаточно. Итак, направление эволюции определяется двумя взаимосвязанными факторами естественного отбора, под влиянием которых сформировалось повсеместно наблюдаемое явление: по мере приближения к нулевым широтам цвет кожи местных жителей становится все более темным. Эти обусловленные солнцем изменения цвета кожи и волос вырабатывались из поколения в поколение в процессе длительной эволюции. Те факторы естественного отбора, которые определяют темный цвет кожи, очевидно, повлияли на генетический код жителей Африки. Если же покинуть Африку и отправиться путешествовать по обширным территориям Евразии, спускаясь из Скандинавии, на севере Европы, и Сибири, на севере Азии, к землям Италии и Юго-Восточной Азии, на которых расположены южные оконечности обоих континентов, можно наблюдать, как постепенно меняется цвет кожи и волос местных жителей. Подобно африканцам, другие темнокожие народы живут преимущественно в солнечных тропических странах. Вполне очевидно, что мы столкнемся и с исключительными случаями, но они будут представлены потомками тех людей, которые в недавнем прошлом покинули родину и поселились в южных странах, их кожа будет еще слишком светла для этих широт (или наоборот, слишком темна), что и естественно, ведь процесс эволюции длится многие поколения. Таким примером-исключением может служить Австралия, которая заселена потомками недавних эмигрантов, поэтому жители этой страны по большей части до сих пор остаются «бледнолицыми». В Австралии, как нигде на земном шаре, высок процент людей, страдающих раком кожи, но и здесь уже начался постепенный процесс эволюции, который, по всей вероятности, ведет к тому, что потомки выходцев из Европы приобретут естественный для этих широт темный цвет кожи. И наоборот, первые переселенцы, отправившиеся с юга на север в страны Северной Европы и Азии, выглядели неестественно темными среди местных жителей, но со временем кожа их потомков стала более бледной. Однако оставим на время Австралию. Дело в том, что в какой бы точке Земли мы ни оказались, цвет кожи местных жителей будет связан с местным уровнем интенсивности ультрафиолетового излучения (рис. 5.1)[260]. 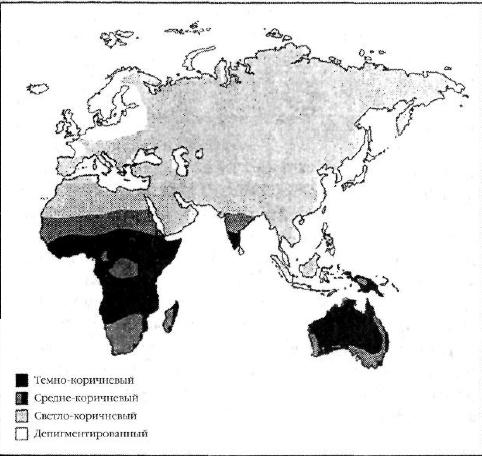 Рис. 5. 1 Поскольку эта особенность эволюционного развития постоянно оказывает влияние на человека во все эпохи и на всех широтах, наш цвет кожи способен поведать скорее о том, где жили наши предки в последние 10—20 тысяч лет, нежели об их генетической дивергенции за предыдущие 60 тысяч лет. Поэтому цвет кожи у современного человека имеет весьма ограниченную ценность как источник, позволяющий проследить древние маршруты миграции разных ветвей человечества после его исхода из Африки. Другими словами, цвет кожи — это далеко не самый надежный индикатор для изучения доисторической эры развития человечества, скажем, между 70 000 и 10 000 гг. до н.э. Может показаться странным, что цвет кожи столь изменчив, поскольку именно он является самым заметным и решающим среди всех «расовых» признаков. Ключом к дискриминационным установкам в поведении человека являются не логические доводы, а стремление отыскать какую-нибудь легко узнаваемую деталь или предлог, позволяющие объединить представителей своей группы и «отсечь» членов соперничающих групп. Различие между ними может носить самый тривиальный характер. Мы регулярно выдумываем все более абсурдные различия, например, различия между всевозможными религиозными сектами, чтобы оправдать преследования или истребление наших соседей. Изменение формы и черт лица Другие, более существенные изменения, такие, например, как форма нашего лица, обусловлены различиями лицевых костей черепа. Эти лицевые кости претерпели существенные изменения у жителей Восточной, Юго-Восточной и Южной Азии, что является отражением достаточно длительного периода, прошедшего после разделения населения этих регионов на разные ветви. На всей территории Восточной Азии мы встречаем монголоидный тип (см. табл. 16) с характерной для него дополнительной складкой, так называемым эпикантусом[261], защищающим верхнее веко, а также широкими скулами и черепом. Этот тип часто делится на два подтипа: северных монголоидов и южных монголоидов (см. рис 5.2), причем для последних характерен менее выраженный эпикантус; в том числе южно-китайский и более смуглокожий монголоидный типы, распространенные в Юго-Восточной Азии[262]. В Южной Азии, в частности — в Индии, можно встретить множество самых разнообразных этнических групп. Большинство индусов, несмотря на темно-смуглый цвет кожи, по телосложению и прочим антропологическим параметрам гораздо ближе к европейцам и жителям Ближнего Востока, чем к обитателям Южной Азии. Европейцев, имеющих длинный узкий череп, круглые глаза и бледную кожу, иногда называют представителями кавказского типа. Чем дальше мы продвигаемся на север Индии и Пакистана, тем ближе жители этих районов к «кавказоидам» — обитателям Леванта и ближневосточного региона. Напротив, в южной Индии преобладают представители смуглокожего, курчавого и круглоглазого этнического типа. В восточной Индии, Ассаме и Непале чаще встречаются люди с более выраженными монголоидными чертами внешности. В главе 4 мы рассматривали другой тип обитателей Азии, которые, возможно, являются потомками племен, селившихся вдоль древнего прибрежного пути в Австралию. По всему побережью Индийского океана, в Пакистане и Индии, на Андаманских и Филиппинских островах, а также в Малайзии сохранились отдельные, так называемые аборигенные, этнические группы, которые по внешнему облику более напоминают африканцев и жителей Новой. Гвинеи. У них очень темный цвет кожи и характерные курчавые волосы. Эти этнические меньшинства принято называть соответственно негроидами, негритосами и меланезийцами. Хотя эти аборигенные группы и составляют ничтожно малое меньшинство среди прочих великих и малых народов азиатского Старого Света, о которых я уже упоминал, они, по мнению многих ученых, в физическом и антропологическом отношении гораздо ближе к первым племенам-выходцам из Африки, чем к другим народам Азии. Это сходство убедительно свидетельствует о тех огромных изменениях, которые претерпело подавляющее большинство этносов Азии. Изучение строения зубов у тех же аборигенных групп населения, которое я использовал в качестве генетических индикаторов (глава 4), позволило получить целый ряд антропологических аргументов в пользу этой точки зрения. Другим подтверждением того, что эти аборигенные группы представляют собой реликты древних обитателей побережья и островов, является тот факт, что ископаемые останки черепов, найденные в Европе, Восточной и Юго-Восточной Азии и восходящие к эпохе до последнего Ледникового периода, по форме нельзя отнести ни к кавказоидному, ни к монголоидному типу. Вместо этого мы видим формы, близкие к древнейшим черепам африканского и ближневосточного типа, возраст которых составляет 100 тысяч лет[263]. Древние люди были более рослыми и крепкими Известно ли нам, как могла выглядеть первая группа эмигрантов-переселенцев из Африки? Проще всего ответить, что нет. Наиболее естественное и правдоподобное предположение о том, что они были темнокожими и имели курчавые волосы, основано на внешности современных африканцев[264], хотя весьма маловероятно, что африканцы, жившие 80—100 тысяч лет тому назад, выглядели, как их современные потомки. И тем не менее ряд ключей к этой проблеме все же существует. До нас дошло несколько ископаемых черепов людей современного (в анатомическом смысле) типа, возраст которых — около 100 тысяч лет. Один из них, который часто называют типичным примером черепов того периода, найден в районе первого неудавшегося исхода из Африки на Левант (Ближневосточный регион). Он был назван Скхул 5 — по названию пещеры, где он был обнаружен. В тех же местах был найден еще ряд аналогичных черепов, так что Скхул 5 — всего лишь наиболее хорошо сохранившийся из них. Череп типа Скхул использовался для создания реконструкции «лица Евы» (см. Фото 17) в нашем документальном фильме «Истинная Ева». Черепам людей анатомически современного типа того периода присуща широкая вариативность, но они обладают несколькими общими особенностями, которые претерпели изменения в последующих поколениях, в том числе и у современных африканцев. Наиболее характерными из них являются крупный рост и «крепость» черепа. В числе факторов, обусловивших крепость, — крупные, покатые черепа, толстые кости и тяжелые надбровные дуги, резко отличающие древних людей от современного человека. В отличие от «крепкого» типа, так называемый «стройный» вариант используется для описания людей с плоским, высоким черепом, вертикальным лбом, более плавными надбровными дугами и сравнительно более тонкими костями конечностей и черепа. В числе других наследственных признаков черепов эпохи, предшествовавшей южному исходу, можно назвать относительную узость черепа (меньшее расстояние от виска до виска по сравнению с расстоянием между лбом и затылком) и более широкую верхнюю часть лица[265].  Фото 17. Так выглядела Ева выходцев из Африки. Реконструкция, использованная в документальном фильме «Путь из Эдема»/«Истинная Ева». Реконструкция воссоздана по одному из наиболее хорошо сохранившихся черепов эпохи исхода на Левант. Ее черты отражают толстокостный тип, типичный для той эпохи, относительно узкий череп и широкие скулы и лоб. Крупный рост и крепость скелета тесно связаны друг с другом (это в особенности относится к черепу). Это ставит перед нами серьезную проблему, когда мы пытаемся воспользоваться костями для реконструкции человека доисторического периода, поскольку наиболее заметным изменением со времен исхода народов из Африки явилось все более нарастающее снижение роста и крепости телосложения у всех народов мира, в том числе — африканцев. Хотя некоторые из устойчивых изменений носят, по-видимому, генетический характер, наиболее резко выраженное снижение роста и крепости телосложения у человека за последние 10 тысяч лет обусловлены, как ни парадоксально это звучит, особенностями питания, а не генетическими факторами. Почему мы стали более мелкими? Древние охотники и собиратели вели суровую, исполненную опасностей жизнь, и поэтому плотность населения была крайне низкой, но зато их рацион был очень богат белками и минеральными веществами, а также всевозможными овощами и фруктами. Наоборот, хотя земледельцы за последние 8000 лет сумели добиться более высокой урожайности зерновых с гектара и имели большие семьи в результате раннего отнятия детей от груди, их рацион отличался однообразием. Их пища была богата углеводами, но скудна белками, витаминами и кальцием. Раннее отнятие от груди и переход на пищу, богатую углеводами и бедную белками, приводил к задержкам в росте, которые сохранялись и во взрослом возрасте. Следовательно, хотя с развитием земледелия численность населения значительно возросла, рост и физическая выносливость детей и взрослых резко уменьшились. Этот эффект особенно заметен у народов, чей рацион изобилует рисом, но крайне скуден животными белками, примером чему могут служить народы Дальнего Востока. Сегодня мы имеем возможность наблюдать, как сказывается обратное изменение этой культурно-обусловленной формы недоедания. Когда уроженцы стран Восточной Азии, основой питания которых на родине был рис, эмигрировали в США и переходили на совершенно иной — высококалорийный, богатый белками — рацион питания, дети бывших азиатов уже через два поколения почти догоняли в росте американских детей. Известно, что после Второй мировой войны дети жителей Европы и Америки благодаря высококалорийному питанию превосходят своих родителей в росте и физических данных[266]. Таким образом, если крепость телосложения до некоторой степени зависит от пропорций тела, а средние пропорции жителей различных регионов, в свою очередь, зависят от их сегодняшнего рациона питания, получается, что роль этих двух важнейших составляющих нашей внешности в формировании различий между неафриканскими народами за последние 70—80 тысяч лет заметно снизилась. Кроме того, многие негативные изменения крепости, пропорций тела, а также внешних данных современных людей со времени исхода из Африки вполне могут быть выправлены с помощью полноценного рациона и здорового образа жизни. Именно этим объясняется типично «неандертальский» облик некоторых современных «звезд» жестких контактных видов спорта. Со временем формируются устойчивые изменения, среди которых особенно выделяются два фактора: изменения черепа и зубов. Прежде всего надо отметить, что уменьшение размеров черепа в разных регионах было обусловлено различными факторами. Изменения черепа Хотя аборигены Австралии и в меньшей степени некоторые племена Новой Гвинеи тоже претерпели негативные изменения — уменьшение роста и пропорций тела, — они, оказывается, сохранили значительно большую крепость скелета, чем практически все остальные народы мира. Уменьшение величины черепа у австралоидов затронуло как его высоту, так и ширину. Аборигенов Австралии иногда называют лучшим примером сохранения архаических наследственных форм, однако еще более полноценными «консерваторами» являются жители Новой Гвинеи, что объясняется целым рядом причин. Что касается аборигенов Австралии, как в прошлом, так и в настоящем, то некоторые из присущих им черт остаются необъяснимыми. Одна из них заключается в том, что древнейшие ископаемые останки черепов, найденные в Австралии, принадлежали к «стройному», а не к «крепкому» типу. Другая причина, позволяющая считать черепа австралоидов несколько «изменившимися» по сравнению с архаическим типом, заключается в том, что у большинства аборигенов Австралии в наши дни волосы не вьющиеся, а курчавые[267]. В наши дни существует всего лишь одна этническая группа, сохранившая такой же уровень крепости черепа, как у австралийцев и жителей Новой Гвинеи. Это — племя тьерра дель фуэган в Южной Америке, сегодня уже практически вымершее. В Японии найден другой череп «крепкого» типа: это знаменитые ископаемые черепа народности минатогава с острова Окинава, субтропического японского острова, датируемые — судя по высоте — последним Ледниковым периодом (см. Фото 18). Эти черепа весьма близки по форме к черепам пренеолитической этнической группы йомонов, также живших на Японских островах, — тех самых йомонов, которые считаются предками айнов, современного коренного населения Северной Японии[268]. У предков европейцев и жителей Среднего Востока (Средней Азии) наблюдались ограниченные и характеризующиеся широкой вариативностью изменения длины и крепости черепа. К ним относятся длинноголовость (дилихоцефалия), а также значительное снижение крепости черепа, что помогает объяснить такую деталь, как мои собственные насупленные брови[269].  Фото 18. Череп Минатогава I с о.Окинава, датируемый последним ледниковым максимумом, вполне может считаться останками древнейших предков аборигенов Японии — айнов. Наиболее же значительные изменения отмечены у народов, которых в наши дни принято называть монголоидами. У них уменьшение величины черепа приняло характер заметного сокращения расстояния между лбом и затылком при сохранении ширины и высоты черепа. Другое важное изменение, характерное для монголоидной расы, — подчеркнуто плоский тип лица. Эта особенность наиболее четко прослеживается на черепах людей эпохи неолита, живших на восточном берегу озера Байкал в Сибири, а также по всей Монголии (рис. 5.2). Как было установлено, выраженно плоский характер лица не является принципиально новой чертой в общем диапазоне вариативности форм человеческого черепа. Плоское лицо — правда, в менее выраженном виде — характерно для некоторых ранних черепов древнейших жителей Африки, для современных охотников и собирателей народности хойсан, а также для достопочтенного отца-основателя[270] Южно-Африканской Республики после падения режима апартеида. Подобные изменения привели к появлению и распространению высокого черепа с тонкими чертами (брацицефалического типа), преобладающего в наши дни в Восточной Азии, что имеет под собой уже четко сложившуюся генетическую основу. Жители Восточной Азии относятся к числу обладателей самых «стройных» черепов среди современного человечества[271]. Когда же и где возникли и получили распространение физические изменения, характерные для современного облика представителей монголоидной расы? Конкретная дата появления в Китае первых людей современного типа вызывает споры, однако это, скорее всего, произошло около 70 тысяч лет назад. При этом важно помнить, что первые обитатели Китая современного типа не имели ничего общего с внешностью монголоидного типа и что немногочисленные ископаемые останки черепов, относящиеся к эпохе до последнего Ледникового периода и найденные в равнинном Китае, возраст которых составляет около 20 тысяч лет, относятся к архическому «крепкому» типу. Фактически же возраст самых древних в мире останков бесспорно монголоидного типа составляет всего 10 тысяч лет. Хотя эти останки были найдены на Дальнем Востоке, самые древние останки человека современного типа с монголоидными чертами были найдены далеко на западе, в урочище Афонтова Гора II на берегах реки Енисей на юге Сибири, что к западу от озера Байкал. Поскольку эта изолированная находка вызывает целый ряд вопросов и других черепов, относящихся к доледниковому периоду, в Южной Сибири не найдено, все, что можно сказать об этом единичном ископаемом черепе без ссылки на другие источники, — это то, что локусом изменений в направлении монголоидного типа не обязательно был Китай[272]. 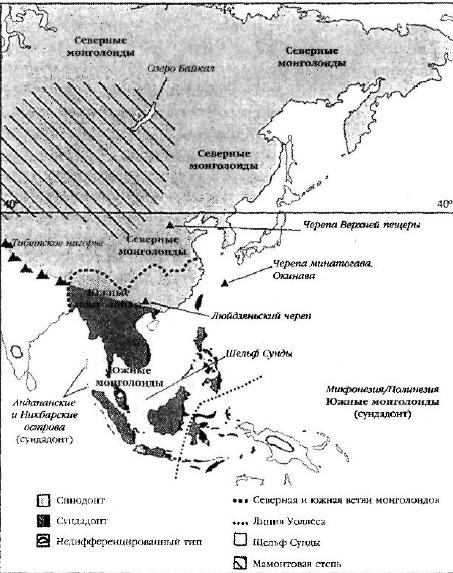 Рис. 5.2 Эволюция зубов Другими, не столь заметными, но не менее важными изменениями явились изменения в форме зубов. Сохранение некоторых архаических особенностей скелета у современных европеоидов стало причиной проведения целого ряда детальных дентологических исследований (рис. 5.3), которые показали, что европейцев тоже можно поставить в один ряд с обитателями прибрежной полосы Индийского океана (включая жителей Южной Индии, негритосов племени семанг (Малайя) и коренное население Новой Гвинеи), сохранившими куда больше архаических африканских особенностей строения зубов и черепа, чем представители любых других народов за пределами Африки. Эта заметная близость в строении зубов у европейцев и других неафриканских народов региона Индийского океана, которым присущи «наименее выраженные изменения», видимо, является следствием одного южного исхода. На мой взгляд, это свидетельство — еще один гвоздь в гроб (если таковой еще нужен) расистских стереотипов, объявляющих европейцев высшей расой, а аборигенов — примитивными низшими существами[273]. К этим «недифференцированным по дентальным признакам обитателям прибрежной полосы» с точки зрения дентального анализа весьма близки представители обширной группы народов. В ее состав входят аборигены Юго-Восточной Азии, принадлежащие к несемангскому (т.е. не связанному с негритосами) типу, в частности — малайцы, представители коренного населения Малайского полуострова, различные архаические этнические группы Тихоокеанского побережья, такие, как аборигены-айны в Японии, а также многочисленные полинезийские племена. Все эти группы сохранили те или иные признаки сходства с представителями ранней формы строения зубов — сундадонтами, название которых происходит от названия геологического района в Юго-Восточной Азии — шельфа Сунды (Сундаленд), где, как предполагается, возник и сформировался этот тип. Сундадонты не встречаются далее к западу вдоль прибрежной полосы, например, в Индии. Монголоидные черты выделяются здесь в особый тип, который можно классифицировать как южных монголоидов. Народности минотагава (палеолит) и йомоны в Японии, которые могут считаться предками современных айну (айнов), также имеют в строении зубов черты, сближающие их с сундадонтами Юго-Восточной Азии и племенами островов Тихого океана[274]. 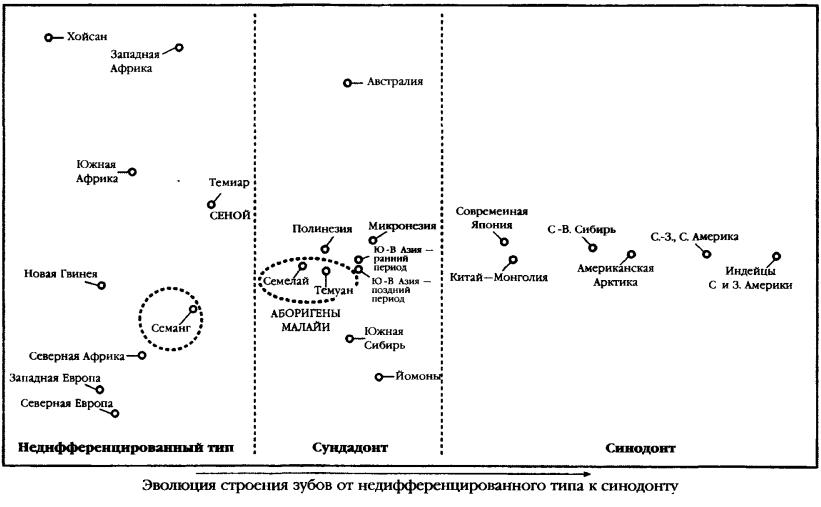 Рис. 5.3 Особое место в этом ряду (см. рис. 5.3) принадлежит аборигенам Австралии, которые, в отличие от своих соседей из Новой Гвинеи, сохраняют некоторые черты, свойственные сундадонту. Эта тенденция свидетельствует о возможности промежуточного смешения[275]. Вторая из двух крупнейших дентологических групп Евразии, помимо сундадонтов, — это синодонты (букв. «китаезубые»). Для этнических групп синодонтов характерно сохранение некоторых особенностей строения зубов, присутствовавших у сундадонтов, а также общее снижение величины зубов. Синодонтность присуща всем северным монголоидам, обитающим в Центральной Азии, причем ее характерные свойства становятся более выраженными при продвижении на север, что относится и к жителям Америки, переселившимся туда из Азии через Берингов пролив (см рис 5.2). Для Юго-Восточной Азии синодонтность даже в наше время является нехарактерной. Прибегнув к сильному упрощению, можно сказать, что сундадонтность отражает древнейшие изменения в строении зубов по сравнению с собирателями на прибрежной полосе — выходцами из Африки, и разнообразные комбинации этих черт можно встретить у представителей южной ветви монголоидов, обитающих в Юго-Восточной Азии, Полинезии и Микронезии, и в меньшей мере у некоторых немонголоидных народностей, таких, как йомоны в Японии и изолированные этнические группы жителей Андаманских островов. Синодонтность представляет собой следующий этап дивергенции в том же морфологическом направлении, что и сундадонтность. Она встречается у северной ветви монголоидов, включая и большинство китайцев, и в более выраженном и гипертрофированном варианте — у жителей Северо-Восточной Сибири и коренных жителей Америки[276]. Загадка происхождения монголоидов При изучении формы черепа, строения зубов или других генетических индикаторов нельзя не заметить, что среди всех этнических групп за пределами Африки наиболее сильные изменения со времени исхода затронули именно монголоидную расу. Эта дифференциация по сравнению с древнейшим архаическим прототипом и утрата внутригруппового разнообразия заметно усиливается по мере продвижения на крайний Северо-Восток Азии и далее — в американскую Арктику. Главный вопрос, остающийся неясным в наши дни, — как, когда и где началось активное формирование черт, выделяющих представителей монголоидной расы по сравнению с другими народами Евразии. И это — не праздный схоластический вопрос, занимающий лишь академические круги. В конце концов, именно монголоиды — наиболее многочисленная и распространенная раса на земном шаре. Ее представители появились в Америке еще до последнего Ледникового периода, примерно 20 тысяч лет тому назад (см главу 7), однако что касается равнинных районов Китая, то там этот тип появился лишь после того, как была перевернута последняя страница в истории последнего оледенения. Краткое резюме этой сложной и запутанной истории в нашем изложении выглядит так: 1. Древнейшие обитатели Азии (которые были также и предками европейцев) внешне выглядели практически так же, как и африканцы той эпохи, и имели сходное строение зубов. 2. Эти древнейшие жители Южной Азии, обладавшие «крепким» черепом, впервые появились около 80 тысяч лет тому назад и расселились по всей прибрежной полосе Южной Азии, от побережья Аравийского полуострова через Индию в Юго-Восточную Азию, выйдя к берегам Новой Гвинеи в конце дуги островов на юго-западе Тихого океана. Современные жители районов вдоль этого маршрута расселения сохранили некоторые архаические черты строения зубов, характерные для их далеких предков. 3. Из Индокитая представители этих недифференцированных типов расселились на север, в Китай, в период между 35 и 70 тысячами лет тому назад обогнув западное побережье Тихого океана. 4. В некий исторический момент еще до последнего Ледникового периода среди представителей «премонголоидной расы», живших в Юго-Восточной Азии, сформировался новый комплекс особенностей строения зубов. Эти новые типы строения зубов впоследствии в несколько видоизмененной форме распространились на северо-восток, вдоль побережья Тихого океана, и дальше — в Японию (их носители стали предками современных йомонов и айнов), а также на юго-восток — в Полинезию. (См. рис 5.2 ) 5. Второй основной комплекс признаков строения зубов, синодонтность, по всей видимости, является более поздним и происходит от сундадонтности. Эволюция от сундадонтности к синодонтности и связанные с ней промежуточные формы не выявлены ни в Юго-Восточной Азии, ни на западной окраине Тихого океана, ни среди йомонов или айнов в Японии. Однако степень синодонтности постепенно нарастает по мере продвижения на север у современного монголоидного населения континентальных районов Восточной Азии, достигая своего крайнего выражения у коренных жителей Америки. Таким образом, получается, что источником и предшественником синодонтности была сундадонтность, возникшая в южных районах. В связи с отсутствием ископаемых окаменелостей признаки синодонтности на черепах старше 10 тысяч лет, найденных в Китае и на Японских островах, обнаружить не удалось. 6. Монголоидной форме черепа присущи тонкие «удлиненные» кости, характерное плоское лицо и уменьшение длины. Подчеркнуто плоский нос обнаружен у одного из черепов, относящихся к эпохе до последнего Ледникового периода, найденном в районе возле озера Байкал. Плоский нос наиболее выражен у черепов в этом районе. 7. Как и синодонтность, признаки монголоидной расы на китайских черепах, относящихся к эпохе до последнего Ледникового периода, не выявлены. В более поздние времена они получили широкое распространение в Северной и Восточной Азии и считаются характерной особенностью синодонтов, то есть, другими словами, северной ветви монголоидов, а также большинства коренных народностей Америки. Таким образом, нарастающие изменения формы черепа протекали параллельно с переходом от сундадонтности к синодонтности в строении зубов. Если предками синодонтов действительно были выходцы с юга — сундадонты, то по крайней мере некоторые черты физического облика, характерные для северной ветви монголоидов, также восходят к южным предкам. Если же предки первых типичных монголоидов северной ветви были выходцами с юга, это означает, что они имели общего предка с такими народами, как малайцы — коренные обитатели Малайского полуострова, которые до сих пор живут там. Как мы увидим далее, существуют вполне убедительные свидетельства того, что по крайней мере некоторые основные генетические цепочки Восточной и Северо-Восточной Азии берут свое начало из Индокитая, сохраняя расовые черты и признаки, которые и в наши дни можно встретить у коренных представителей южной ветви монголоидов в этом регионе. В этой связи сделать паузу и рассмотреть различные частные случаи этой модели миграции «юг — север», представляющей собой противоположность модели «север — юг», что прослеживается по изменениям строения зубов и костей. Из двух основных азиатских типов строения зубов, ассоциируемых с представителями монголоидной расы, сундадонтность, по всей видимости, возникла первоначально в Юго-Восточной Азии, представляя собой древнейший тип изменения строения зубов, который и в наши дни встречается у некоторых реликтовых архаических этнических групп немонголоидного происхождения. Синодонтность же, напротив, является самым распространенным типом строения зубов во всей Восточной Азии и Америке, однако она не встречается у представителей коренного населения Юго-Восточной Азии. Последний аргумент говорит в пользу версии направления миграции «юг—север». А как же быть с гипотезой о миграции с севера на юг? Если она справедлива и корни происхождения южной ветви монголоидов именно таковы, мы вправе были бы ожидать массового вторжения носителей синодонтных признаков с севера на юг, в Юго-Восточную Азию. Однако на самом деле ничего подобного не обнаружено. Простой анализ уровня изменений также опровергает теорию о том, что специфические типы строения зубов привели сначала к возникновению сундадонтности на юге, а затем трансформировались в синодонтность, когда носители монголоидных признаков направились в Центральную Азию. Что касается эволюции и усиления признаков сундадонтности, что повлекло за собой формирование синодонтного типа, некоторые признаки монголоидной расы, например характерная плоскостность лица, более выражены у северной ветви монголоидов, чем у южных монголоидов. Таким образом, какими бы ни были движущие силы эволюции, эти специфические изменения должны были накапливаться где-то еще дальше к северу, возможно, в глубине континентальных районов Азии. Ископаемые останки (или, точнее сказать, их отсутствие) свидетельствуют о том, что район «дальнейшего развития» монголоидных признаков находился отнюдь не в Китае, Японии, Юго-Восточной Азии или Индии, а где-то в Центральной Азии к северу от Гималаев, например, в Монголии или в районе озера Байкал в Южной Сибири. Более того, на юге Сибири существует особая этническая группа, среди представителей которой, кстати сказать, резко отличающихся от соседних народностей, встречается немало носителей сундадонтных признаков, что свидетельствует о сохранении в этих местах архаического расового типа (см. рис. 5.3). Почему представители монголоидной расы стали выглядеть иначе? Существует целый ряд теорий, объясняющих, как и где начали формироваться особенности скелета монголоидов, обусловившие их внешние отличия. Первое, что необходимо сказать об этих теориях, — это то, что все они носят откровенно спекулятивный и даже спорный характер. Теории, рассматривающие вопрос «где», имеют, как я уже отмечал, чисто гипотетический характер, что объясняется практически полным отсутствием ископаемых останков представителей монголоидной расы. Недостоверность же теорий, отвечающих на вопрос «как», обусловлена тем, что сегодня не существует реальных средств, позволяющих связывать те или иные изменения скелета с конкретными генами-«контролерами». Впрочем, это не мешает нам хотя бы вкратце познакомиться с такими теориями. К сожалению, как это часто случается в научных дебатах, противоречия между сторонами приводят к возникновению настоящей пропасти между ними, тогда как истина обычно находится где-то посередине. Что касается теорий о происхождении, то нам предлагается выбор между южным и северным локусами прародины предков монголоидов, а если говорить о теориях, объясняющих, как могли произойти подобные изменения, нам приходится обращаться к адаптирующей физической эволюции, при которой возможны либо изолированное развитие, либо дрейф (миграция) генов. (Впрочем, между двумя предполагаемыми механизмами изменений существует определенная связь, поскольку адаптационные[277] теории рассматривают именно холодную северную зону в качестве среды, где имела место адаптация.) Географическое деление монголоидов на северян и южан становится еще более заметным при рассмотрении физических (и генетических) факторов, как старых, так и выявленных совсем недавно, свидетельствующих о том, что единая некогда раса действительно разделилась на две ветви — северных и южных монголоидов. Изучение динамики роста у детей в Китае показало, например, что дети жителей Гонконга (Южный Китай), несмотря на то что питались они гораздо лучше детей из континентального Китая, заметно уступали в росте детям из Пекина (Северный Китай). Тот факт, что дети из Гонконга, питающиеся лучше своих сверстников из Пекина, значительно уступают им в росте даже спустя несколько поколений после экономического бума в Гонконге, свидетельствует о том, что между севером и югом сложились устойчивые генетические различия[278]. Являются ли расовые отличия монголоидов результатом дрейфа генов или целенаправленного отбора? Основную альтернативу традиционным гипотезам о происхождении монголоидов, рассматривающим проблему, «как» они возникли, сформулировать достаточно несложно. Наиболее простой ответ на этот вопрос (рассматриваемый в последней главе) сводится к тому, что немногочисленная группа-основатель, изолированная от прочих этнических групп, самостоятельно эволюционирует, приближаясь к определенному генетическому или физическому типу. Комбинированное воздействие эффектов группы-основателя и накапливания изменений еще более усиливается в результате сексуального отбора. Чтобы упростить этот пример еще более, буквально до абсурда, допустим, что в группе-основателе монголоидной расы, пришедшей с юга, у большинства ее представителей по чистой случайности оказались маленькие носы. В результате подобной групповой самоидентификации и предпочтения, отдаваемому при выборе партнеров лицам с маленькими носами, курносые носы через какое-то время стали основной отличительной чертой внешности членов группы. Это еще более усилило действие эволюционных сил, способствовавших формированию маленьких носов, и у дальних потомков этой группы-основателя носы стали совсем крошечными. В дрейф генов оказались вовлечены и другие факторы, ассоциируемые в генетическом отношении с маленькими носами, например плоские лица. Другая теория, более старая и, возможно, более спорная, заключается в том, что основателям монголоидной расы пришлось адаптироваться к куда более неблагоприятным и сложным условиям новой среды обитания, в данном случае — к холодному и ветреному климату азиатских степей к северу от Гималаев. Эта идея имеет давнюю родословную, но удачнее всех, пожалуй, ее сформулировал американский биолог Дэйл Гатри. Гатри увлеченно описывал среду обитания, которую он называл Мамонтовыми степями Азии эпохи палеолита. Мамонтовая степь представляла собой необъятные пространства, поросшие высокими травами; сердцем этой степи был обширный район, расположенный к северу от 35-й параллели, Гималаев и Тибетского нагорья. Этот комплекс факторов низкотемпературной среды обитания, известный биологам под названием «палеолитический биом», прекратил существование около 10 тысяч лет тому назад, являя собой парадоксальный результат глобального потепления, а вместе с ним навсегда исчезли и множество крупных и мелких видов флоры и фауны[279]. По мнению Гатри, в южной части «сердца» Мамонтовой степи очень рано появились люди, пришедшие сюда из Синьцзяна, Монголии и окрестностей озера Байкал в Сибири. Сегодня в этих местах раскинулись обширные площади пустынь. В эпоху расцвета великая Мамонтовая степь простиралась от побережья Атлантики на западе до острова Хоккайдо на востоке. Это была самая крупная травянистая саванна, когда-либо существовавшая на нашей планете, и в ней спокойно паслись неисчислимые стада крупных травоядных. Небольшие стада некоторых из этих травоядных дожили до наших дней в труднодоступных районах. К числу этих животных принадлежат дикая лошадь, северный олень, мускусный бык и антилопа сайгак, Что же касается более крупных видов, таких, как волосатый мамонт, степной бизон и волосатый носорог, то они давно вымерли. Понятно, что Мамонтовая степь была поистине раем для охотников, которые научились преодолевать холода и сильные ветры. Гатри пишет, что люди эпохи палеолита, охотившиеся на травоядных животных в этой степи, предпочитали селиться вдоль южной ее границы, проходившей примерно по 40-й параллели, где было больше островков леса, дававших людям приют и укрытие (см. рис. 5.2). Нет никаких сомнений, что люди, жившие в столь холодных климатических условиях, нуждались в теплой одежде. Гатри и другие исследователи высказывают предположение, что выжить в суровых условиях будущим монголоидам помог целый ряд факторов физической адаптации. В их числе — двойное верхнее веко, или уже упоминавшаяся складка эпикантус, позволявшая уменьшить потери тепла через глаза, более толстый слой подкожного жира вокруг глаз и на щеках, скулах и подбородке; маленький нос, снижающий риск обморожения; более округлая форма головы и особое строение черепа, позволяющее организму терять меньше тепла; более плотное и приземистое туловище с относительно короткими конечностями; более равномерное распределение жира, что, вместе с меньшей длиной конечностей, обеспечивало снижение на 10% отношение поверхности тела к его объему; бледная, близкая по тону к европейцам кожа, обеспечивающая поглощение солнечных лучей и выработку достаточного количества витамина D для защиты от рахита; и, наконец, повышенная чувствительность кровеносных сосудов в конечностях, позволяющая избежать обморожения. Лично я, несмотря на то что целый ряд аргументов Гатри представляется мне весьма вероятными, нахожу их перечень чрезмерно обширным, да и аргумент о величине носа не кажется мне убедительным. Ведь в Европе, где климатические условия практически такие же, как современные ее жители, так и неандертальцы использовали и используют большие носы как своего рода кондиционеры для согревания и увлажнения воздуха перед тем, как он попадет в весьма чувствительные бронхи и легкие. Это представляется куда более убедительным аргументом в пользу появления крупных носов, и подобные гипотезы высказывались как минимум дважды независимо друг от друга. В самом деле, разве может маленький нос быть столь же приспособлен к жизни в холодном климате? Между тем именно у эскимосов и в меньшей степени — чукчей, жителей крайнего северо-востока Сибири, развился нос, который принято называть «приплюснутым»: крошечный нос, буквально вдавленный в плоское лицо[280]. Дальнейший процесс адаптации монголоидов к условиям жизни на Крайнем Севере свидетельствует, что крошечный нос оказался не слишком эффективным «кондиционером», и климат Арктики потребовал его дальнейшей эволюции. Наиболее достоверным (под словом «достоверность» я имею в виду обоснованность теоретических построений, опирающихся на реальные факты) в рассуждениях Гатри мне представляется место, где он утверждает, что наиболее активно адаптация протекала на севере, где у северной ветви монголоидов наблюдается сравнительно бледная кожа, а самые выраженные изменения внешнего облика, в частности появление особенно плоского лица, наблюдаются в окрестностях озера Байкал как в глубокой древности, так и у современных коренных жителей этого региона[281]. Гатри высказал ряд предположений относительно того, сколь долгим мог быть процесс развития адаптационных изменений и когда он начался. Он вполне обоснованно полагает, что наибольшие изменения подобного рода вызвал последний Ледниковый период, достигший своей кульминации около 20 тысяч лет тому назад, создав так называемый стресс холода, но говорит, что сам процесс мог начаться как минимум 40 тысяч лет тому назад. Аргументы в поддержку столь раннего начала сводятся к тому, что, как мы уже говорили, накапливание изменений и признаков плоского лица началось у будущих монголоидов, живших к западу от озера Байкал, задолго до начала Ледникового периода (см. рис. 5.2). Хотя я должен признаться, что теоретические построения Гатри относительно эволюционного развития представляются мне весьма соблазнительными, меня тревожит проблема отсутствия реальных, материальных доказательств подобного рода — ископаемых останков человека. Кроме того, нерешенным остается и вопрос о том, каким образом у представителей южной ветви монголоидов, живущих в тропиках Юго-Восточной Азии, могли появиться эти черты внешности, если эти люди, как принято считать, являются потомками оседлого коренного населения. Если весь комплекс признаков, общих для северной и южной ветвей монголоидов, в первую очередь обусловлен адаптацией к холодным климатическим условиям, в таком случае, согласно теории миграции с юга на север, он должен был бы отсутствовать у представителей южной ветви монголоидов. Позволю себе привести почти анекдотический пример. Моя дражайшая жена, предки которой были уроженцами южного побережья Китая и которая по классификации, используемой мною в этой книге, относится к южным монголоидам, переносит холод куда хуже, чем я, человек, предки которого были южанами! Если же теории о миграции с юга на север и адаптации к холодному климату соответствуют действительности, нам пришлось бы признать, что первоначальные физические изменения могли явиться результатом действия некоего иного эволюционного механизма, чем простая адаптация к холоду, и что некоторые из этих изменений впоследствии были заметно усилены под воздействием холода. Неотения у человека Мне представляется весьма убедительным предложенное Гатри объяснение воздействия эволюционных факторов на севере, но не менее правдоподобным выглядит и продвижение с юга на север, подтверждаемое генетическими данными и особенностями строения зубов. Между тем существует и другой эволюционный феномен, способный связать воедино два этих фактора. Много лет назад видный палеонтолог Стивен Джэй Гоулд выдвинул интересную гипотезу, заключающуюся в том, что комплекс анатомических изменений, характерных для монголоидов, можно объяснить феноменом неотении — явления, при котором инфантильная или детская форма тела сохраняется во взрослом возрасте. Неотения у человека является одним из наиболее простых объяснений того, как и почему у людей за последние несколько миллионов лет так быстро развился непропорционально большой мозг. Относительно большой мозг и обращенность черепа вперед на позвоночнике, исчезновение волосяного покрова на всем теле, характерные для человека, наблюдаются у внутриутробных, неродившихся детенышей шимпанзе. Гоулд обратил внимание на постепенное усиление признаков неотении у монголоидов и даже предложил для этого явления особый термин — «педоморфия»[282]. Такой механизм, по всей видимости, включает в себя всего один ген-«контролер», и поэтому подобный эффект может быть достигнут за относительно короткий промежуток эволюции. Он же вполне может послужить объяснением появления совершенно нелогичного курносого носа и исчезновения основной части волосяного покрова на лице. Существует несколько эволюционных механизмов, в результате которых особи с выраженным педоморфизмом могут со временем произвести на свет сравнительно больше потомства, что объясняется дрейфом генов или направленным генетическим отбором, причем оба этих фактора не являются взаимоисключающими. Один из этих механизмов может быть обусловлен генами экономичности, использующимися в холодной степи (сокращение излишней мышечной массы, уменьшение объема зубов, более тонкие кости, меньший рос и физические данные). Это соответствует избирательно-адаптационной модели эволюции монголоидов, которая могла включать в себя уже существующие признаки, возникшие и оформившиеся у сундадонтов, которые мигрировали с юга на север вдоль побережья Тихого океана. Точно так же женщины с педоморфными признаками могли считаться более привлекательными, а это уже аргументы в пользу гипотезы полового подбора и дрейфа генов. И хотя дальнейшие дрейф или подбор могли протекать на севере, сам комплекс признаков вполне мог быть занесен туда с юга. Я склонен полагать, что в эволюции монголоидного типа могли сыграть свою роль оба этих признака. Признаки южной ветви монголоидов первоначально могли сформироваться в результате полового отбора и/или дрейфа генов. Затем, когда некоторые из носителей этих признаков переселились в Центральную Азию, у них началось «дальнейшее развитие «темы», обусловленное теми самыми эволюционными факторами, о которых так увлеченно писал Дэйл Гатри. В этом смысле могло существовать две основных «домашних» платформы для развития признаков монголоидной расы. Эта кабинетная теория не нашла широкой научной поддержки, поскольку генетика пока что находится на пороге определения того, какие именно гены контролируют рост и развитие телосложения, не касаясь вопроса о выявлении основных различий внутри нашего собственного вида. С другой стороны, генетический анализ, вероятно, смог бы оказать немалую помощь в определении направления миграции. Четыре пути в Центральную Азию Как и всегда случается в подобного рода походах и экспедициях, осуществляемых как современными людьми, так и их далекими предками, на решение о выборе дальнейшего пути вновь прибывшими переселенцами в Азии повлияли два фактора — географические особенности местоположения и климат. Правила выбора места всегда были достаточно просты: старайся селиться там, где есть вода и выпадает достаточно дождей, а во время миграции избегай пустынь и высоких горных хребтов и следуй вдоль берегов рек. Мы уже располагаем рядом свидетельств того, что маршрут собирателей на прибрежной полосе вдоль побережья Индийского океана на пути в Австралию действительно был наиболее легким и удобным. Чем же это объяснить? Ведь на самом деле маршрут был далеко не из простых; дело в том, что поначалу через каждые несколько сотен километров нашим первопроходцам-мигрантам приходилось переправляться вброд через устья больших рек. Но раз они совершали подобные переправы на пути в Австралию, им, естественно, приходилось проделывать то же самое и вдоль всего побережья Восточной Азии. И на берегу очередной великой реки у переселенцев была возможность повернуть влево и направиться в глубь материка, питаясь дарами речных вод и следуя вверх по ее течению. Как заметил Марко Поло, один из первых европейцев-путешественников, побывавший в Восточной Азии, пустыни и горы представляют собой труднопреодолимые преграды для всякого, кто стремится проникнуть в Центральную Азию. За исключением немногочисленных перевалов, путь туда возможен только вдоль долин, образуемых руслами больших рек. Как мы знаем, первый успешный исход из Африки, увлекший далеких предков всех неафриканских народов, имел место примерно 75 тысяч лет тому назад. Продвигаясь вдоль прибрежной полосы, они могли достаточно рано проникнуть в Китай и Японию. На этом пути они, по всей видимости, обошли стороной весь Центральноазиатский регион. Однако они в любой момент своего дальнего странствия могли свернуть влево и двинуться в глубь материка вверх по течению одной из великих рек. На севере Индии, где в небо вздымаются заснеженные пики Гималаев, дальнейший путь был сопряжен с серьезными трудностями. Гигантские складки горных хребтов, возникшие в результате грандиозной тектонической подвижки, которая произошла в Азии в глубокой древности, высятся по обе стороны высочайших вершин Гималаев, охватывая Непал и Тибет. Высокие горные хребты, практически всюду достигающие высоты 3000 м, наглухо закрывают доступ в Центральную Азию со стороны побережья Индийского океана на протяжении более 6500 км от Афганистана на западе до Ченьду, Китай, на востоке. Там, на востоке, эта гряда расстилается как скомканный ковер, образуя целый ряд хребтов, протянувшихся с севера на юг еще на 2500 км — от начала Шелкового пути в северном Китае до Таиланда на юге. Шелковый путь, получивший широкую известность на Западе благодаря тому же Марко Поло, представлял собой длинный торговый маршрут[283], протянувшийся к северу от Гималаев и проходящий параллельно им. Этот маршрут связывал Восток с Западом. Он проходил через всю Центральную Азию, практически совпадая с южной границей сердца Мамонтовой степи, о которой писал Гатри. Шелковый путь и в наши дни, как и в древности, остается одной из немногих артерий, связывающих Китай с Западом, если не считать протяженный путь, ведущий на юг, к Сингапуру. На восток — по Шелковому пути от западной оконечности Гималаев. В наши дни Шелковый путь проходит по южной и северной границам пустыни Такламакан в Синьцзяне. В эпоху палеолита на месте нынешней пустыни лежали травянистые степи, а еще дальше к северу находился целый ряд водных путей, в том числе — реки Тарим и Джунгария, служившие легким и удобным путем для охотников и собирателей из западных районов Центральной Азии, таких, как Таджикистан, Узбекистан, Киргизстан и Казахстан, в Синьцзян[284] и Монголию. Этими же водными артериями могли воспользоваться и древнейшие люди, стремившиеся проникнуть в Центральную Азию[285]. А теперь обратимся к реалиям Каменного века. Первые люди могли проникнуть в Центральную Азию с юга вверх по течению Инда, протяженность которого достигает 8000 км, и достичь западных районов Китая у западной оконечности Шелкового пути. Предположим, что речь идет о первых мигрантах, оказавшихся в Индии. Важнейшей задачей, стоявшей перед ними после подъема вверх по течению Инда, было преодоление горных хребтов на севере Индии и Пакистана. Хребты эти простирались далеко на запад, до Афганистана. Обойти эти горы с запада и двигаться дальше на север, через территорию современного Афганистана, было делом крайне сложным, если не сказать — невозможным, ибо, не говоря уже о невероятно длинном обходном пути на запад, мигрантам пришлось бы идти практически через пустыни. Марко Поло пересек эти пустыни. Он отправился в путь из Ормуза в устье Персидского залива и, пройдя через Афганистан, оказался в Кашмире, после чего, миновав горный перевал, попал в Китай и направился в Кашгар. Оттуда он, уже по Шелковому пути, двинулся в самое сердце бывшей Мамонтовой степи. Впрочем, Марко Поло мог попасть в Кашмир и гораздо более легким путем. Немного восточнее, неподалеку от побережья современного Пакистана, великий Инд описывает петлю и поворачивает на север, до того места, где начинается водный путь, по которому можно попасть в Кашмир. Другой, тоже относительно равнинный, путь в Центральную Азию через верховья Инда ведет через Хайберский перевал в Кабул, а оттуда — в Узбекистан и Казахстан и далее на восток, в Синьцзян. На запад — по Шелковому пути из Китая. На всем протяжении истории Шелковый путь служил единственным маршрутом, ведущим из Китая на запад — в Центральную Азию. Таким образом, альтернативный путь, ведущий в Мамонтовую степь, мог пролегать от тихоокеанского побережья Восточной Азии. Передовые отряды древнейших мигрантов, двигавшихся вдоль береговой линии, могли пройти по всему побережью Китая и, оказавшись в северном Китае, повернуть на запад и по Шелковому пути проникнуть в Монголию, Синьцзян и Южную Сибирь. На север — из Бирмы в Тибет. Этот, третий путь в Мамонтовую степь, не слишком часто используемый в наши дни в качестве торговой артерии, ведет к восточной оконечности Гималаев. Восточная окраина Гималаев состоит из множества отрогов и складок, образовавшихся в том месте, где Индийская материковая плита уперлась в Азиатскую, что вызвало тектонические подвижки. Как видно на карте (рис. 5.4), эти складки представляют собой как бы естественные каналы для большинства великих рек Южной и Юго-Восточной Азии. Реками этими, если двигаться с запада на восток, являются Брахмапутра, протекающая в Бангладеш, Сальвин, несущая свои воды по территории Бирмы, Меконг, текущий по землям Вьетнама, и Янцзы, которая течет в южном Китае. Эти четыре великие реки, вытекающие из северо-восточных отрогов Тибета, несут свои воды практически параллельно друг другу на протяжении свыше 150 км, а расстояние, разделяющее их на этом участке, составляет буквально считаные километры. Последняя из четырех великих рек, Янцзы, берет свое начало в горах северо-восточного Тибета у северной окраины плато, возле самой границы Мамонтовой степи. Я упомянул об этих руслах великих рек не потому, что многие торговцы и в наши дни активно пользуются этими водными артериями, а потому, что они позволяют попасть в Тибет[286], Монголию и Центральную Азию через одно из четырех устий рек, которые находятся на большом расстоянии друг от друга на побережье Юго-Восточной и Восточной Азии и впадают в Индийский и Тихий океаны. На восток — из России. Наконец, следует назвать еще один, северный маршрут миграции с запада на восток, в Восточную Азию: через земли азиатских районов России, так называемый русский Алтай. Наиболее удобный сухопутный путь из района Алтая в Центральную Азию во время самого мягкого в климатическом отношении периода Позднего Каменного века (30—50 тысяч лет назад) пролегал непосредственно через степи. Продвигаясь на восток через территорию Южной Сибири по многочисленным озерам и рекам, существовавшим в те времена, наши древние мигранты-первопроходцы могли достичь района озера Байкал по маршруту, пролегающему через Синьцзян и Монголию. В те времена степи на этих землях были покрыты густыми высокими травами. Тем не менее для того, чтобы проделать этот путь, мигрантам первым делом надо было попасть на Алтай. Как мы увидим, примерно 40 тысяч лет назад они действительно достигли Алтая и окрестностей озера Байкал в Южной Сибири. Итак, теперь мы знаем, что в древности существовало четыре потенциальных маршрута, ведущих в Центральную Азию: три из них вели от побережья Индийского и Тихого океанов (с запада, юга и востока), а один брал свое начало на территории современной России (с северо-запада). 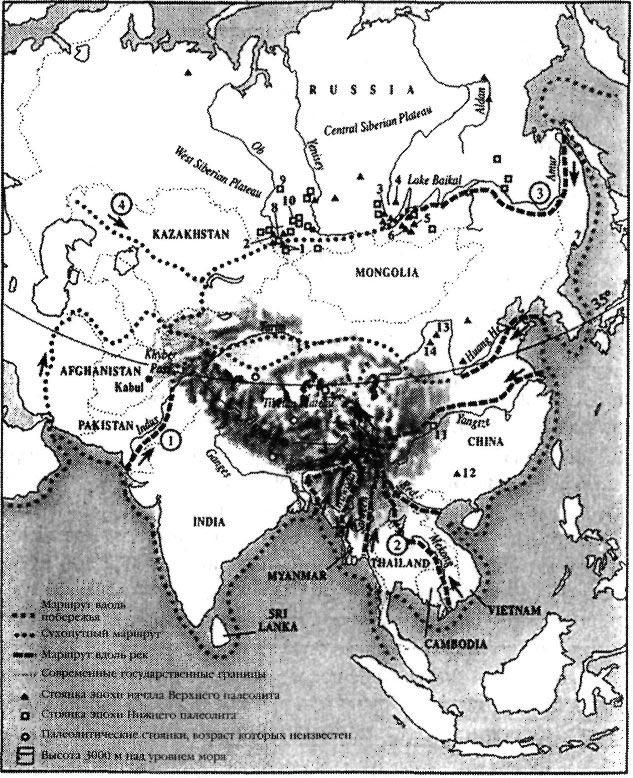 Рис. 5.4 В самой же Центральной Азии существовало три параллельных маршрута вдоль водных артерий, соединявших восток и запад, — маршрута, по которым могли пройти первопроходцы древности. Два из них — южные: через Синьцзян и Монголию, и один — северный через Южную Сибирь. Северный маршрут был доступен только в периоды потепления в эпоху палеолита, то есть от 30 до 50 тысяч лет назад, во время так называемых климатических пауз (см главу 3). Когда люди современного типа впервые отправились в странствия по степям? На сегодняшний день наиболее ясные и достоверные археологические свидетельства присутствия людей современного типа к северу от Гималаев более 40 тысяч лет тому назад нам удалось обнаружить именно вдоль четвертого и самого северного из этих коридоров, ведущих в степи. Южная Сибирь находится буквально у самого сердца Мамонтовой степи, которое, согласно трудам Гатри, находилось в Синьнцзяне и Монголии. Впечатляющие результаты последних археологических раскопок в пещерах Алтая — Усть-Каракол и Кара-Бом (см .рис 5.4) — со всей ясностью показывают, что новая технология обработки камня эпохи Верхнего палеолита появилась в этих местах около 43 тысяч лет назад. Эти датировки находок на Алтае весьма близки по возрасту к другим находкам, сделанным далее к востоку отсюда, в районе села Макарово на западном берегу озера Байкал, возраст которых составляет около 39 тысяч лет. В период между 30 и 35 тысячами лет тому назад существовал целый ряд подобных стоянок. Варварина Гора и Толбага на восточном берегу озера Байкал и Малая Сия, далее к северу, у берегов Енисея. Это означает, что переход к Верхнему палеолиту на всей территории Южной Сибири произошел как минимум не позже аналогичного перехода в Европе[287]. Не так давно бельгийский археолог Марсель Отте заявил, что центр зарождения новой технологии обработки камня эпохи Верхнего палеолита находился на севере нагорья Загрос (на территории нынешних Ирана и Ирака), откуда эта технология впоследствии распространилась как на северо-запад — в Левант и Европу, так и на северо-восток — в Центральную Азию и на Алтай (Россия), предположительно — по тому пути, который описывал еще Марко Поло, или через Кавказ[288]. Способны ли каменные орудия поведать о том, какие племена первыми появились в Центральной Азии? Появление новой технологии обработки камня эпохи Верхнего палеолита принято считать решающим свидетельством проникновения человека современного типа в Европу (на Балканы и далее на запад). Она же считается своего рода индикатором перехода от неандертальца, для которого характерны орудия эпохи Среднего палеолита. Как известно, этот переход произошел несколько сотен тысяч лет назад (см. главу 3). Это можно считать свидетельством параллельного прибытия людей современного типа, обладателей новой технологии обработки камня эпохи Верхнего палеолита, с юга, с нагорья Загрос, на Алтай, что произошло около 43 тысяч лет назад[289]. Впрочем, если обладание этой новой технологией не являлось этаким sine qua nоn[290] для появления людей современного типа в Азии, то вполне вероятно, что первые люди могли проникнуть на Алтай и в более ранний период. Правда, надо признать, что к датировке времени прибытия первых людей в любые районы Азии необходимо подходить с особой осторожностью. Во-первых, хотя технология обработки камня, возникшая в эпоху Верхнего палеолита, действительно свидетельствует о присутствий человека современного типа в Европе, а также на территории Леванта и Юго-Восточной Азии, она была отнюдь не единственным техническим знанием, которым пользовались люди современного типа, так что свидетельства о ее применении в тех или иных районах нельзя использовать в качестве опоры для датировки появления первых людей. Дело в том, что эпоха Верхнего палеолита явилась, образно говоря, собранием целого ряда новшеств, первое из которых, согласно новейшим археологическим данным, появилось около 45—50 тысяч лет тому назад, и его принято ассоциировать с Северной и Западной Евразией. Первые люди современного типа использовали целый ряд разнообразных «старых» технологий и навыков, включая традиционные способы изготовления орудий, унаследованные от Среднего палеолита и характерные для неандертальцев. Находки в пещерах Алтая (Россия), в частности — в пещере Кара-Бом, свидетельствуют о наличии традиционных техник Среднего палеолита в слоях, залегающих ниже типичных ауриньякских слоев Верхнего палеолита и датируемых 62—72 тысячами лет назад. Эти нижние слои трудно отнести к деятельности людей современного или архаического типа (в частности, неандертальцев), поскольку они с равным основанием могли быть оставлены и теми и другими, и к тому же в них не сохранилось ископаемых останков и артефактов, позволяющих уточнить датировку. Ученые никогда не находили останки неандертальцев так далеко на севере — на Алтае, хотя на юго-западе, например в Тешикташе (Узбекистан, Центральная Азия), подобные останки — не редкость. Ключевой вопрос здесь состоит в том, отражают ли более ранние слои Среднего палеолита постоянное накапливание технических навыков, позволившее совершить прорыв в технологии изготовления орудий, или культура Верхнего палеолита была занесена откуда-то извне. Последнее утверждение представляется более реальным, по крайней мере, в том, что касается ауриньякской технологии. Таким образом, сохраняется реальная вероятность того, что слои эпохи Среднего палеолита, возраст которых составляет 60—70 тысяч лет, найденные в пещере Кара-Бом, представляют собой свидетельства присутствия человека современного типа на Алтае в еще более раннюю эпоху[291]. Вторая причина, побуждающая проявлять осторожность в датировке, заключается в том, что вплоть до недавнего времени хронология палеолита в Синьцзяне, Тибете и южной Монголии была практически не изучена. Дело здесь не в отсутствии останков и артефактов эпохи палеолита, а в том, что многие из них не поддаются датировке. Один из недавних отчетов о раскопках в Монголии утверждает, что возраст традиционных технических навыков Среднего палеолита, по-видимому, составляет 60 тысяч лет. В другом отчете говорится, что на Тибете существовали две раздельные микролитические традиции эпохи Верхнего палеолита: северная и южная, связанные с жителями и навыками обработки каменных орудий соответственно Северного и Южного Китая. Датировка была не слишком достоверной, поскольку многие находки были обнаружены на поверхности (находки с поверхности, естественно, вырваны из исторического контекста и реального соположения слоев, необходимых для археологической датировки)[292]. Таким образом, мы располагаем весьма убедительными археологическим свидетельствами того, что технология эпохи Позднего палеолита распространялась и северным путем, через всю Южную Сибирь, и начало этому процессу было положено на Алтае (Россия) примерно 43 тысячи лет тому назад. С другой стороны, более чем вероятно, что в Центральной Азии и на юге, в сердце Мамонтовой степи, уже в древности активно селились люди современного типа, однако реальных ископаемых останков, подтверждающих это, пока не обнаружено. Как мы вскоре увидим, данные генетических исследований, полученные в Северной Азии, служат весомым аргументом в пользу того, что люди современного типа появились в Северо-Западной Азии около 40 тысяч лет назад. Люди эпохи Верхнего палеолита, жившие в Северной Азии, во многом близки к представителям богатой граветтийской культуры, возникшей в Мамонтовой степи далеко на западе — в Центральной и Восточной Европе, о чем свидетельствуют находки в урочище Мальта неподалеку от озера Байкал, возраст которых — 23 тысячи лет (см. табл. 19)[293]. Генетические данные Я проанализировал многие археологические материалы, свидетельствующие о присутствии первых людей современного типа в Центральной Азии, а также на побережье двух океанов — у «входов» в единственные коридоры, ведущие в Центральную Азию, к Шелковому пути и в самое сердце Мамонтовой степи. Эти трое «ворот» находились на северо-западе Азии, в Индокитае и в Северо-Восточном Китае. Что же касается генетических данных, то их я упоминал очень скупо, и на то были свои причины. Структура ветвей митохондриевой ДНК и Y-маркеров является куда лучшими индикаторами древнейших миграций молекул, чем средством датировки или идентификации так называемых рас или этнических групп. Таким образом, куда надежнее — разумеется, там, где это возможно, — прослеживать маршруты предполагаемых миграций с учетом географических, археологических и климатических данных, прежде чем выяснять, в какой мере эти результаты подтверждаются генетическими данными. Деление на север и юг в Восточной Азии Я уже упоминал в главе 2 о том, что после первичного рассеяния народов — выходцев из Африки были постепенно заселены все регионы и земли Старого Света и юго-западная часть Тихого океана и что вплоть до наступления последнего великого Ледникового периода (около 20 тысяч лет тому назад) межрегиональная миграция генов была крайне незначительной или отсутствовала совсем. И Азия, несмотря на всю свою богатую историю переселения народов, отнюдь не явилась исключением из этого правила. Некоторые крупные генетические ветви на азиатском континенте стали известны генетикам задолго до появления понятия о митохондриевой ДНК и Y-маркерах. Как и следовало ожидать, регионы распространения кавказоидного типа в Западной Евразии располагались поблизости друг от друга, а некоторые из них «накладывались» на Индийский субконтинент. При этом имело место выраженное отличие от Восточной Евразии, население которой, в свою очередь, подразделялось на северян и южан, что соответствовало северной и южной ветвям монголоидов. Если говорить о происхождении будущих народов, мы должны провести четкую границу между Юго-Восточной Азией (включая южное побережье Китая: провинции Гуандун, Гуаньси и Фудзян) и Северо-Восточной Азией (включая Северный Китай, восточное побережье Китая, Японию, Корею, Монголию, Тибет и некоторые районы Сибири), причем одна из групп населения, живущая на крайнем северовостоке Азии, в этническом отношении занимает промежуточное положение между кавказоидами и северными монголоидами[294]. Подобные исследования мало что дают для выявления локусов формирования народов, но, несмотря на это, показывают, что даже в крупных этносах, таких, как так называемый этнос хань в Китае, прослеживаются выраженные региональные варианты, тяготеющие к дифференциации по принципу север — юг. Эти исследования помогли также развеять целый ряд мифов о генетических истоках происхождения китайцев. Так, например, широко бытовало мнение, что за последние несколько тысяч лет существования императорской власти в Китае[295] имела место массовая экспансия ханьцев с севера на юг, следствием которой якобы были вытеснение, замещение и ассимиляция коренного населения этих регионов. Однако современные исторические исследования показывают, что народности Южного Китая, исходя в первую очередь из прагматических политических соображений, стали называть себя ханьцами совсем недавно — всего каких-нибудь несколько веков назад[296]. Это свидетельствует о том, что большинство миграций и переселений народных масс в Китае носило скорее легендарный, нежели реальный характер, и поддерживает точку зрения о том, что состав региональных этнических групп населения в Китае был куда более устойчивым, чем это считалось прежде. Другой подход к решению вопроса о том, сформировались ли монголоиды как раса на севере или на юге, — это анализ вариативности. Исходя из основных принципов учения Дарвина, несколько видов (или в данном случае новая этническая группа, развившаяся на основе небольшой группы-основателя) должны были иметь общий центр происхождения, сохраняя при этом широкий спектр вариантов (в том числе и закрепленных генетическими изменениями) в пределах такой группы. Сегодня, когда доступен генетический анализ, у южной ветви монголоидов неизменно наблюдается гораздо более широкий спектр вариантов, чем у северной. Изучение изменений Y-хромосомы также позволяет считать родиной монголоидов именно южные районы[297]. Несмотря на всю свою пользу, применение подобного упрощенного подхода к вариативности имеет два потенциальных недостатка. Во-первых, ни один из генных путей или маркеров, использованных в исследованиях, нельзя идентифицировать с признаками «монголоидности»: останки людей отождествляют с монголоидами просто по месту находки, и поэтому подобный аргумент, если применять его с достаточной последовательностью, рискует оказаться в замкнутом круге одних и тех же посылок и выводов. Во-вторых, Северная Азия, в отличие от тропической Юго-Восточной Азии, во время последнего Ледникового периода сильно пострадала от депопуляции (сокращения численности населения), массового вымирания людей и генного дрейфа, так что низкая вариативность, наблюдаемая сегодня на севере, отнюдь не обязательно отражает первоначальную этническую картину. Но, пожалуй, наиболее радикальная гипотеза, описывающая влияние последнего Ледникового периода на население Центральной и Северной Азии, сводится к тому, что в древности весь обширный азиатский регион, простирающийся к северу от Гималаев, был совершенно безлюдным, так что нынешние жители этих мест являются потомками выходцев из Южного Китая и Юго-Восточной Азии, а также Европы. Это, разумеется, означает, что мы так никогда и не узнаем, каким был первоначальный генетический комплекс древнейших жителей Северной Азии. Эта точка зрения представляется не слишком убедительной. Во-первых, детальные палеоклиматологические[298] исследования показывают, что во время Ледникового периода в Центральной Азии существовали обширные пространства степной тундры, способные обеспечить всем необходимым охотников. Это подтверждается археологическими находками, отражающими основные виды занятий человека во время максимальной фазы последнего Ледникового периода (см. главу 6). Дэйл Гатри решительно выступает против гипотезы о полном отсутствии населения на юге, в сердце Мамонтовой степи. Во-вторых, как мы увидим ниже, подобная картина не позволяет объяснить наличие генетического многообразия и поливариантности на юго-западе Центральной Азии (в Узбекистане и Киргизстане) или резко выраженных географических различий между представителями разных ветвей генетического древа в Северной и Южной Азии. В частности, существует целая группа генетических линий, единых для Северной Европы, Северной Азии и Северной Америки, однако не имеющих ничего общего с Южным Китаем или Юго-Восточной Азией[299]. Схема развития генетического древа Как и в случае с историей исхода из Африки, ответ на интересующий нас вопрос станет более ясным, если мы посмотрим на маркеры Адама и Евы. Большим достоинством построения древа митохондриевой ДНК и Y-хромосомы является то, что они позволяют проследить ветви и даже побеги, соответствующие отдельным молекулам, применительно к каждому конкретному району. Эту увлекательную научную «игру» в гены и районы их распространения можно назвать филогеографией. В принципе она заключается в соединении разрозненных региональных побегов в крупные ветви и размещении последних на генетическом древе. Хотя правила предельно просты, их применение может оказаться достаточно сложным. Чтобы проследить векторы миграции из одного региона в другой, нам необходимо найти ветви-источники в обоих регионах и новый «побег» в искомом районе, который отсутствует на ветви прародины. Для прародины с многообразными векторами миграции в самых разных направлениях нам необходимо выявить общие ветви, имеющие разные «побеги» в других районах. Пожалуй, один из наиболее удачных примеров подобной методики был описан в работе видного итальянского генетика Антонио Торрони и его коллег, опубликованной в 1993 г. 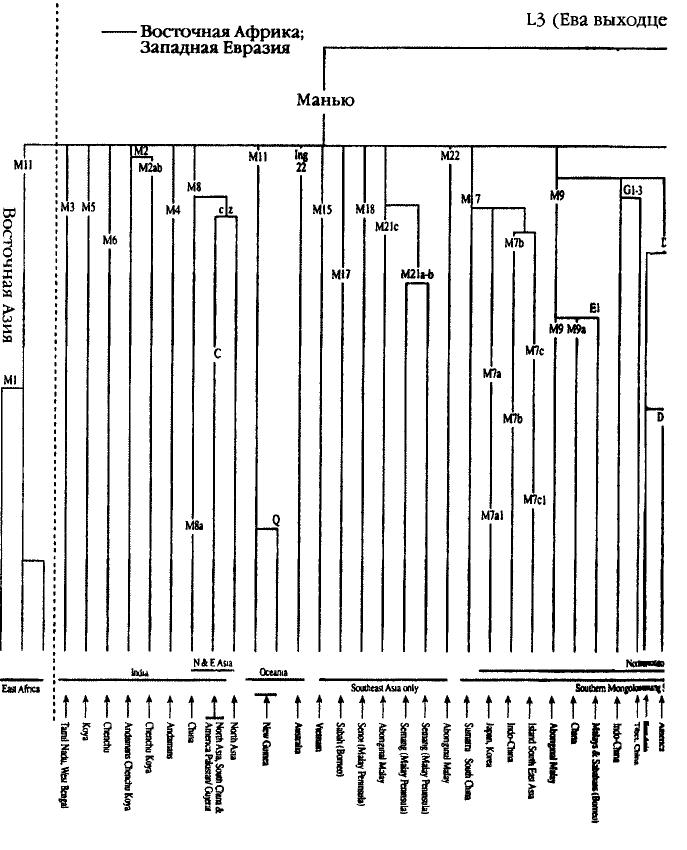 Правая часть 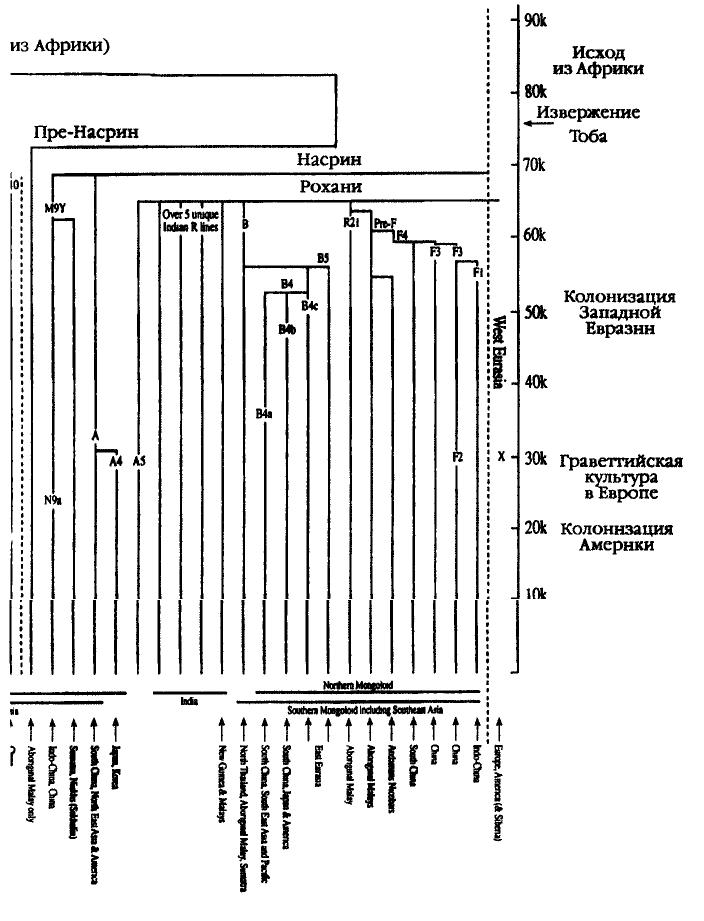 Левая часть Рис 5.5 Эти исследователи выявили четыре основные линии митохондриевой ДНК этнической группы-родоначальницы коренного населения Америки, пометив их буквами А, В, С и D. Истоки каждой из этих буйно разросшихся ветвей, давших обильные «побеги» в Америке, можно возвести к аналогичным ветвям-прародителям А, В, С и D в Азии (см. главы 6 и 7). Мы можем применить этот же подход и к гораздо более сложной проблеме — идентификации прародины монголоидов в Азии. Взяв за начало митохондриевую ДНК Восточной Азии, мы можем заметить, что на сегодня там присутствуют девять четко сформировавшихся ветвей (гаплогрупп; см. рис. 5.5), которые можно обозначить буквами A, D, С, D, E, F, G, X и Z. За исключением сестринских групп С и Z, ни одна из этих групп не может быть выведена на основе другой. Каждая из них более или менее уверенно выстраивается на основе двух евразийских линий-родоначальниц — Манью или Насрин. Недавно были описаны еще несколько второстепенных групп (М7/М10 и N9), однако их существование никак не сказывается на значимости аргументации. (Более подробно о филогенезе и источниках по распределению гаплогрупп митохондриевой ДНК в Восточной Азии см. примечание[300].) Из девяти наиболее хорошо изученных восточно-азиатских ветвей митохондриевой ДНК три (С, X и Z, см. рис. 5.9) прослеживаются только к северу от условной линии «запад — восток», проведенной ниже Гималаев, тогда как остальные шесть (А, В, D, E, F и G) были обнаружены у обеих — как северной, так и южной — ветвей монголоидов. На первый взгляд может показаться, что эти уникальные северные ветви — весомые аргументы в пользу гипотезы о том, что прародина монголоидов находилась к северу от Гималаев, но на самом деле у этого феномена есть более простое объяснение. Дело в том, что линия С и близкородственная ей линия Z, а также загадочная группа X не являются коренными фигурантами древнейшей истории Монголии. Они появились среди северной ветви монголоидов откуда-то извне, вероятнее всего — из Сибири или Центральной Азии. Как мы скоро увидим, в поддержку этой версии существует целый ряд аргументов. Распределение генов и структура шести остальных ветвей генетического древа свидетельствуют о том, что генетическая прародина монголоидов находилась к югу от Гималаев. С особенной ясностью на это указывают древнейшие генетические ветви В и F. Таким образом, мы, по всей видимости, имеем дело сразу с несколькими источниками генов у жителей Юго-Восточной Азии, которые впоследствии распространились на север. Наоборот, среди обитателей Северной Азии мы имеем несколько ветвей, никак не связанных с коренным населением Юго-Восточной Азии. Возвращаясь к той же версии о развитии ветвей, можно отметить, что идея о южной прародине и последующей миграции монголоидов на север подтверждается данными исследования материнских генов и вполне соответствует гипотезе об эволюции сундадонтов в синодонтов (см. главу 5). Чтобы сосредоточить внимание на поисках возможного вторичного северного региона «дальнейшего развития той же темы», нам придется обратиться к обзору географических зон, где присутствуют все шесть ветвей митохондриевой ДНК — А, В, D, E, F и G, — являющихся общими для северной и южной ветвей монголоидов. Таким образом, продвигаясь с запада на восток, мы обнаружим, что в Центральной Азии присутствуют пять основных линий (за исключением G), в Тибете — все шесть, в Монголии — Пять (за исключением Е), в Китае — четыре (за исключением Е и G) и, наконец, в Корее — пять (за исключением[301]). Пожалуй, вряд ли можно считать простым совпадением, что страны с большим числом генетических линий, обычно ассоциируемых с монголоидной расой — Центральная Азия, Тибет, Монголия и Корея, — расположены вдоль 40-й параллели, которая, согласно Гатри, служила своего рода осью Мамонтовой степи (см. рис. 5.6). Основываясь на этих факторах, мы можем задать вопрос о том, каков примерный возраст этих ветвей, расположенных вдоль 40-й параллели. Тоомас Кивисилд установил, что местный возраст четырех из этих ветвей в Монголии весьма близок: ветвь А насчитывает 35 500 лет, В — 40 500 лет или 33 500 лет (см. ниже), D — 44 500 лет и, наконец, F — 42 500 лет. Эти даты значительно «моложе», чем средний возраст тех же линий на юге, в Южном Китае. Но самое важное заключается в том, что они вполне сопоставимы с датами начала активной колонизации Центральной Азии и относятся к эпохе после улучшения климата в Азии, которое произошло примерно 52 тысячи лет назад (см. главу 3) и благодаря которому первые обитатели Азии смогли проникнуть на просторы Мамонтовой степи[302]. И все же: по каким путям и маршрутам люди современного типа проникли в этот обширный регион? Обратившись к анализу двух древнейших линий Юго-Восточной Азии — В и F (идущих от потомков Насрин), которые тянутся из Юго-Восточной Азии в Тибет, Монголию и далее на северо-восток, вдоль побережья Тихого океана, мы видим, что они проникают в глубь материка в Сибири (Северная Азия), а затем останавливаются[303]. В этом отношении распределение этнических групп В и F отражает древнейшее распространение признаков сундадонтности. Как В, так и F достигают наиболее выраженной поливариантности среди туземных этнических групп Таиланда, Вьетнама и Камбоджи, что свидетельствует о том, что их прародиной является Индокитай. Весомым аргументом в поддержку версии о том, что группа F возникла и сформировалась на юге, является вновь идентифицированная гаплогруппа «npe-F», которая распространена среди малайцев — коренных жителей Малайского полуострова (выводы сделаны на основе распространения признаков сундадонтности; см. рис. 5.6). 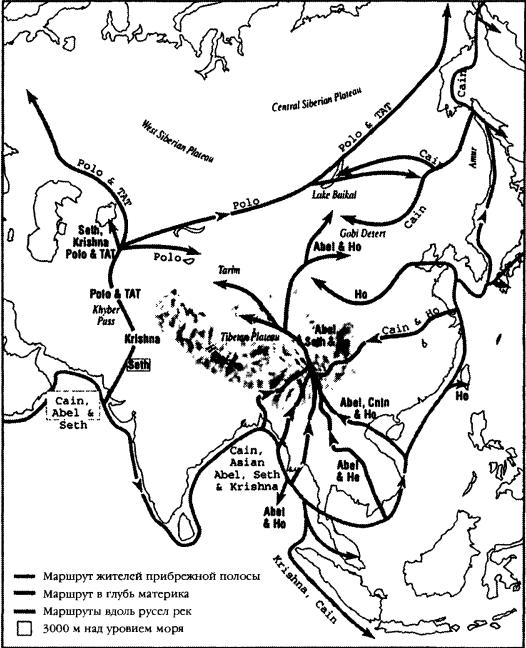 Рис. 5.6 Представители этой группы «пре-F» встречаются в Южном Китае (провинции Юннань и Гуаньси), Индокитае (Таиланд и Вьетнам), на Суматре, а также Андаманских и Никобарских островах, что, по всей вероятности, свидетельствует о древнейших этнических связях с обитателями прибрежной полосы в Юго-Восточной Азии[304]. Располагая сведениями о том, распространились ли представители группы В на запад, проникнув из Китая в Монголию вдоль Шелкового пути, или же двинулись на север из Бирмы через Тибет, мы сможем узнать направление маршрута первой волны колонизации, которая, по всей видимости, имела место около 50 тысяч лет тому назад. Это — немаловажные ключи. Тот же Тоомас Кивисилд установил, что возраст двух основных подгрупп В, проникших в Монголию, составляет 40 500 лет (В1) и 33 500 лет (В2) Это говорит о том, что, хотя в Монголии и части западных районов Центральной Азии оба подтипа В могли появиться еще в эпоху палеолита, более древняя из этих ветвей могла пройти через Тибет, тогда как более поздняя, не исключено, проникла на запад, в Центральную Азию, с побережья Тихого океана, где ее представители сегодня занимают доминирующее положение[305]. Надо упомянуть и о трех линиях — D, Е и G, — частично связанных с Манью, линиях, распространение представителей которых в Восточной Азии увеличивается при продвижении с севера на юг, отражая аналогичное распространение признаков синодонтности. Однако остается нерешенным вопрос о том, выходцами откуда могли быть другие основатели «этнической колонии монголоидов» в Мамонтовой степи (С, X и Z) и каким именно из трех возможных коридоров, ведущих в Мамонтовую степь, они воспользовались. К югу от Гималаев представители этих трех линий Северной Евразии встречаются крайне редко. Ветви-сестры С и Z наиболее распространены в Сибири и Северо-Восточной Азии, тогда как в более южных районах (за исключением Монголии, Тибета и Центральной Азии) их представители почти не встречаются. Линии эти тянутся вдоль всего континуума «восток — запад» через азиатские великие степи, и численность их представителей в Центральной Азии вплоть до Турции весьма значительна. Что касается ветви С, то она простирается даже в Америку через Аляску, однако преимущественное ее распространение на севере Восточной Азии свидетельствует о том, что эти люди не могли прийти в Центральную Азию через Китай. Наоборот, куда более вероятно, что они проникли в Азию откуда-то с запада, причем представители восточной ее ветви были обладателями новой технологии создания орудий, характерной для Позднего палеолита, артефакты которой были обнаружены в пещере Кара-Бом на Алтае (Россия). Возраст их — около 43 тысяч лет. Люди, создавшие их, принадлежали к группе Манью, и их древнейшие предки пришли сюда из Индии и Пакистана, проникнув в долину Инда через Кашмир или Афганистан и обогнув по пути западную оконечность Гималаев (см. рис 5.4) Все эти факты свидетельствуют о том, что мигранты проникли в азиатскую степь где-то 40—50 тысяч лет тому назад из западных районов Центральной Азии (см рис 5.6) Подсчеты Тоомаса Кивисилда, находящиеся в полном соответствии с версией о том, что линии С и 2 распространялись по степи и их носителями были обладатели новой технологии эпохи Верхнего палеолита, показывают, что возраст группы С в Монголии составляет 42 тысячи лет[306]. Последняя из северных линий, линия X, обнаружена только у европейцев и туземных жителей Америки (правда, есть сообщение о находке следов линии X в Южной Сибири), однако связь между Старым и Новым Светом существовала еще как минимум 30 тысяч лет назад. Это свидетельствует о том, что группа X распространялась по необъятным пространствам Центральной и Северо-Восточной Азии вместе с представителями культуры охотников на мамонтов, однако полностью исчезла в Азии во время Последнего Ледникового периода, сохранившись лишь на отдаленных окраинах своего ареала в Америке и Европе (см. главу 7)[307]. Столь сложная картина линий распространения митохондриевой ДНК свидетельствует о том, что монголоиды были выходцами с юга, тогда как жители Центральной Азии происходят от некоей западно-азиатской этнической группы, однако в формирование их родовых признаков внесли свой вклад уроженцы Восточной и Юго-Восточной Азии, мигрировавшие в Центральную и Северо-Восточную Азию. Эти генетические данные являются аргументами в поддержку теории о трех этапах колонизации Центральной Азии с «баз» на побережье Индийского и Тихого океанов 40—50 тысяч лет тому назад. Разграничение и выделение генетических составляющих в этносах и этнических группах Северо-Восточной Азии проясняет вопрос о происхождении ее населения и вполне отвечает концепции о том, что центром происхождения монголоидной расы была Юго-Восточная Азия. История Адама Итак, после того как мы ознакомились с данными генетической структуры ветвей митохондриевой ДНК, изучение истории заселения Азии на общем древе Y-хромосомы и присутствия ряда миграций в Центральную Азию приобретает более достоверный характер. Древо Адама — это во многих отношениях новое понятие в науке. В этом качестве на нем недостает многих и многих «побегов», пышно зеленеющих на Евином древе жизни, а данные для ветвей развития Y-хромосомы весьма и весьма ненадежны. Так ради чего же мне возвращаться к Адаму и лишний раз выслушивать ту же историю, которую куда лучше и подробнее поведала нам Ева? Все дело в том, что Y-хромосома сама по себе является очень четким и достоверным региональным маркером. Выразительный пример этого — история заселения Европы (см. главу 3), где различные патернальные (отцовские) кланы отличались куда более высокой степенью этнической чистоты в рамках Европы, чем материнские кланы Евы. Распространение последних, несмотря на всю их региональную структурированность, остается куда более туманным. То же самое можно сказать и о заселении Восточной Азии. Генеалогическое древо Адама и Евы свидетельствует о том, что и жители Центральной Азии, и аборигены американского континента имеют давние корни, ведущие в Северо-Восточную, Восточную и Юго-Восточную Азию, однако разграничение этнических групп-родоначальниц обитателей Мамонтовой степи гораздо нагляднее прослеживается на Адамовом древе. Чтобы лучше понять этот феномен, давайте вспомним три основные внеафриканские ветви потомков Адама: Авель (YAP или М145), Каин (или RSP4Y) и Сиф (или М89)[308]. (См. рис 5.5.) Линия Каина, или каинитов, — это линия, быстро распространявшаяся на юго-восток вдоль побережья Индийского океана. Каиниты стали мужчинами-родоначальниками этнических групп в Восточной Индонезии, Австралии и Новой Гвинее. Собиратели на прибрежной полосе не ограничивались контактами с населением стран противоположного полушария и продвигались вдоль побережья Индийского и Тихого океанов вплоть до берегов Японии и Кореи, где они создали наиболее ранние свои поселения. Где-то на пути вдоль береговой линии примерно в эпоху Верхнего палеолита, по всей вероятности — ближе к концу своего долгого пути в Японию, у Каина родился сын-азиат, ставший родоначальником многих народов, и его потомки рассеялись по всей Северо-Восточной Азии, а затем по Шелковому пути проникли в Монголию и Центральную Азию. Часть каинитов двинулась дальше на северо-восток и, переправившись через будущий Берингов пролив, оказалась в Америке[309]. В главе 4 мы уже говорили о том, что западные сыновья Авеля приняли активное участие в заселении Европы, Среднего Востока и Африки. Однако существовала и другая, восточно-азиатская ветвь потомков Авеля. Как и каиниты, азиатские потомки Авеля быстро продвигались далеко на восток вдоль побережья двух океанов и достигли берегов Индонезии, а затем двинулись на северо-запад — к острову Тайвань и Японии. Крайней границей их продвижения на север стала Корея. Между тем сегодня в Индии не встретишь потомков азиатской ветви сынов Авеля. Это можно объяснить тем, что они продвигались очень быстро, а их единичные представители в Индии были стерты с лица Земли облаком вулканического пепла, возникшим в результате катастрофического извержения вулкана Тоба, которое произошло примерно 74 тысячи лет тому назад (см. главу 4). Зато в Тибете число азиатской ветви потомков Авеля достаточно велико, что свидетельствует о том, что некоторые из сынов Авеля сумели пережить извержение вулкана Тоба и направились из Восточной Индии вверх по течению великих рек и попали в Юго-Восточной Азии на обширные горные плато, где и обосновались в степях Монголии и на Алтае (Россия)[310]. Таким образом, получается, что оба клана мужчин-мигрантов прибрежной полосы воспользовались двумя восточными коридорами, чтобы проникнуть в степные просторы Центральной Азии (рис. 5.7). Их вклад в колонизацию Старого Света в количественном отношении был ничтожно мал по сравнению с вкладом ветви третьего потомка Адама — Сифа, семя которого было разнесено буквально по всем уголкам неафриканского мира. Его потомки воспользовались всеми тремя путями, чтобы проникнуть в глубинные районы Центральной Азии. По данным последнего исследования, на долю линии потомков Сифа приходится 93% из общего числа 12 127 обследованных жителей островов Азии и Океании, а остальное составляют потомки линий Каина и Авеля. В этой связи важно подчеркнуть, что, подобно дочерям неафриканской Евы Насрин и Манью, сифиты продвигались вдоль береговой линии двух океанов в компании своих собратьев, сынов Каина и Авеля, и, что самое главное, — одновременно с ними. Представители линии Сифа прошли здесь не в более поздние времена и не другим путем, как утверждают некоторые генетики. Доказательством этого служат следы одновременного присутствия потомков всех трех братьев, в том числе — характерно сифские черты внешнего облика у представителей древнейших этнических групп собирателей на прибрежной полосе Индийского океана, в том числе и у жителей Юго-Восточной Азии[311]. 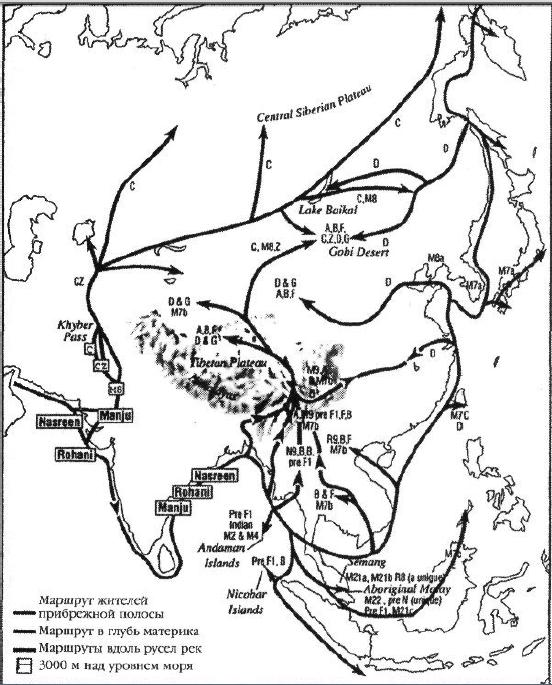 Рис. 5.7 Именно маршрут потомков Сифа подтверждает использование другого, альтернативного пути: того самого северного маршрута, ведущего в Центральную Азию в обход западной оконечности Гималаев (наряду с южной ветвью пути, которая вела в Индокитай и огибала отроги Гималаев с востока). Мы можем даже вычислить, откуда сифиты начали свой поход. Две трети поливариантности типов внешности, присущей этой крупнейшей ветви-родоначальнице евразийских народов, встречается в Индии. На долю самого Сифа пришлась примерно четверть вариантов Y-хромосомы, а на долю его сынов и потомков — большая часть остальных вариантов[312]. Еще примерно 10% потомков Сифа обитают в Центральной Азии, и, следовательно, все четверо его сынов-потомков в первом поколении имели своих представителей в Центральной Азии, что опять-таки говорит о потенциальной возможности прямого расселения их на севере как раз в то самое время, когда Евразию активно заселяли представители других групп с митохондриевой ДНК: С, X и Z. Мы уже знаем, что три сына Сифа внесли основной вклад в колонизацию и заселение большей части Индии, Европы, Среднего Востока и стран Средиземноморья[313]. Однако в этой части нашего повествования нас интересует не столько сам Сиф и три его сына — родоначальника народов Западной Евразии, сколько его четвертый (в генетическом отношении) сын, Кришна, и особенно та роль, которую последний сыграл в формировании отцовских генов многих народов Евразии, не говоря уж о том, что его можно считать отцом практически всех коренных народов Америки. На долю Кришны приходится около 40% типов Y-xpoмосом, обнаруженных на сегодня. Его достаточно широкое распространение в Европе, Азии, Океании и Северной и Южной Америке свидетельствует о том, что он появился на свет в Индии вскоре после первоначального исхода из Африки. Несмотря на достаточно раннее рождение и широкое распространение своей линии, расселение первых «сынов» Кришны носило весьма дискретный характер. Некоторые из них поселились в Пакистане и Индии, другие направились на север, обосновавшись на Леванте и в Центральной Азии, третьих можно встретить только в Меланезии (Новая Гвинея и окружающие острова). Наконец, представители еще одного типа (ТАТ) встречаются только в Центральной Азии и Северо-Восточной Европе[314]. Однако наиболее продуктивными в генетическом отношении сыновьями Кришны — с точки зрения заселения территории — стали два сына Кришны. Каждый из них как бы указывает векторы расселения будущих жителей Евразии и Америки в эпоху палеолита. Одна из этих ветвей, M175, или согласованный тип О, встречается почти исключительно в Восточной и Юго-Восточной Азии. Я назвал бы эту группу Хо, по имени двух выдающихся личностей этого региона: китайского мореплавателя адмирала Чжен Хо (Хэ)[315] и видного деятеля освободительного движения Вьетнама Хо Ши Мина[316]. Если представить, что эта ветвь, как и материнские линии В и F, появилась на свет, когда Кришна, продвигавшийся по прибрежной полосе океанов, прибыл из Индии в Бирму, нетрудно заметить, что Хо, в свою очередь, делится на три ветви. Все три эти ветви сегодня представлены в Китае, Индокитае и Юго-Восточной Азии и отличаются друг от друга лишь степенью расселения на севере. Одна из них осталась в Южном Китае, Индокитае и Юго-Восточной Азии, и на ее долю приходится до 65% аборигенных типов Малайзии. Вторая продолжила вектор расселения до побережья Тихого океана и проникла в Южный Китай, причем наиболее высокая концентрация представителей этой ветви отмечена на о. Тайвань. Третья проникла в глубь континентального Китая и, продвигаясь на север, вдоль побережья, достигла Японии, Кореи и Северо-Восточной Азии, а небольшое число ее представителей проникло по Шелковому пути в Центральную Азию[317]. Характерно, что ни одна из трех этих мужских ветвей, связанных с Восточной и Юго-Восточной Азией, не проникла в Америку (см рис 5.7). В отличие от Восточной и Юго-Восточной Азии, где доминирующая роль принадлежала Хо, для жителей Северной и Центральной Азии основным источником «поставки» Y-хромосом почти исключительно служили потомки другого азиатского «сына» Кришны — Поло (М45). Характер происхождения Поло от Кришны в Индии (рис 5.8) свидетельствует о том, что ранняя ветвь послужила источником Y-маркеров и потомства, представителей которого и сегодня можно встретить среди коренных обитателей региона озера Байкал — кетов и селькупов. Наиболее любопытная в историческом отношении деталь их происхождения заключается в том, что именно Поло — тот самый «сын» Кришны, который дал импульс проникновению представителей этой ветви в Северо-Восточную Европу через Россию, тесно связан с так называемой граветтийской культурой (см главу 3). Мы вправе сопоставить эти материалы с данными распространения этой ветви на Алтае в эпоху Верхнего палеолита — от 23 до 43 тысяч лет тому назад, а также с историей распространения митохондриевых линий в Северной и Центральной Азии. По всей видимости, представители ветви Поло проникли в Центральную Азию из Пакистана примерно 40 тысяч лет назад и расселились на территориях, образующих гигантскую букву «Т». Они продвигались по Мамонтовой степи как на запад, так и на восток. На востоке они достигли берегов озера Байкал и двинулись дальше, в Америку, а на западе проникли сперва в Россию, а затем — в Восточную Европу[318]. 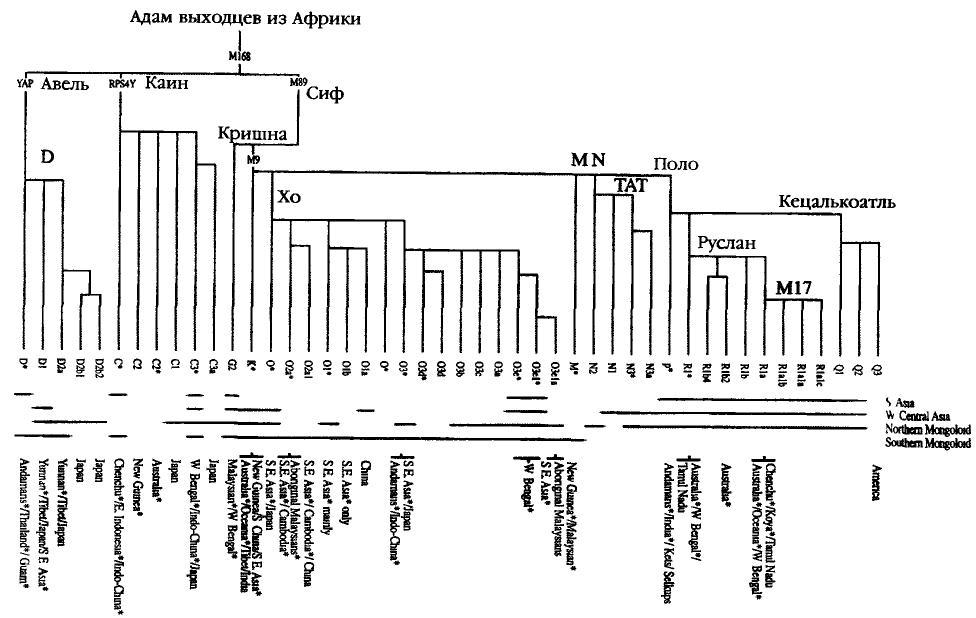 Рис 5.8 Мы располагаем доказательствами столь неординарного «зеркального» распределения митохондриевых Х-маркеров в Северной и Южной Америке, а также в Европе и имеем широкую генетическую базу, свидетельствующую о том, что корни создателей мамонтовой культуры туземцев Америки восходят к жителям Северной Евразии, о чем красноречиво говорит название одного из трудов, опубликованных в 1990-е гг. — «От Костенки до Кловиса: Верхний палеолит и адаптация палео-индийцев»[319]. Представим себе переселенцев, которые двинулись из Индии и Пакистана на север, чтобы основаться на нагорьях Алтая в России. Им пришлось столкнуться с серьезными и негативными климатическими факторами, но усилия их не пропали даром, ибо игра, что называется, стоила свеч. Эта группа-родоначальница населения Северной Азии разделилась на западную и восточную ветви, заняв обширные степи. Те из них, кто направились на восток, впоследствии достигли берегов Америки, а отдавшие предпочтение западу стали прародителями этносов Северной и Западной Европы. Те же, кто решил остаться, во время последнего Ледникового периода оказались у самой его кромки и были вынуждены уйти на юг, в Центральную Азию, а впоследствии совершили повторную экспансию, став теми самыми коренными жителями Сибири и Урала, которых мы видим сегодня на этих территориях. Итак, теперь мы знаем, что древнейшие обитатели прибрежной полосы часто делились на все новые и новые ветви в различных пунктах своего долгого пути вдоль побережья Индийского и Тихого океанов и что примерно 40 тысяч лет тому назад они заселили большую часть Азии и островов Восточного полушария. Первая из ветвей, двинувшаяся из Индии на север, стала родоначальницей охотников эпохи Верхнего палеолита, обитавших в степях Центральной Азии. Последующие же ветви, расселявшиеся вдоль русел великих рек Центральной Азии, дали начало той самой расе, представителей которой мы сегодня называем монголоидами. Свидетельствами о существовании в Азии так называемого монголоидного типа вплоть до максимума Ледникового периода, имевшего место 20 тысяч лет назад, мы не располагаем. Таким образом, эволюционные факторы, обусловившие возникновение этой физической дивергенции, могли действовать и долгое время спустя после прибытия первых выходцев из Центральной Азии в Северную Азию. А это подводит нас к следующему этапу климатической истории нашей планеты — Великому Оледенению. Примечания:2 Richards, M. et al. (2000) «Прослеживание генетических линий предков европейцев в ближневосточном митохондриевом генофонде» American Journal of Human Genetics 67: 1251—76. 3 Эта история описана в книге С. Оппенгеймера — Oppenheimer, S.J. «Эдем на Востоке» (1998, Weidenfeld & Nicolson, London); см. также: Oppenheimer, SJ. and Richards, M. (2001) «Быстрые поезда, тихоходные лодки и предки островитян Полинезии» Science Progress 84(3): 157-81. 25 О дискуссии, посвященной концепции коэволюции Болдуина, см. Deacon, Т. (1997) Symbolic Species (Penguin, London) cc. 322—34. 26 Об эволюции культурных факторов, или мемов, см. Blackmore, S. (1999) «Машина, создающая мемы» (Oxford University Press). 27 Deacon, указ. изд., сс. 214—16. 28 Там же, главы 8 и 9. 29 Там же, сс. 248—50. 30 Теоретически наибольшей «социальной емкостью памяти»: Представители семейства китовых имеют такой же объем мозга, как и человек, но несравненно более крупное туловище. Что касается других животных, то я не раз убеждался, что собаки, имеющие гораздо меньший объем мозга, чем человек, обладают превосходной памятью на индивидуальные запахи. В общей сложности более 300 человек-, см. табл. 2.10 в. Gamble, С. (1999) «Сообщества эпохи палеолита в Европе» Cambridge World Archaeology (Cambridge University Press) с 54. Обмен навыками и материальными ценностями: там же, глава 2. 31 См. дискуссию там же, сс. 53—55. 257 Dunbar, R.I.M. (1992) «Объем неокортекса как фактор, ограничивающий величину стаи у приматов» Journal of Human Evolution 20: 469—93. 258 Jablonski, N. and Chaplin, G. (2000) «Эволюция окраски человеческой кожи» Journal of Human Evolution 59: 57— 106. 259 Остеомаляция — размягчение костей — Прим. перев. 260 Которые определяют темный цвет кожи: т.е. различия в выборе локуса MC1R, что объясняет значительную фенотипическую составляющую выработки меланина — Harding, R. et al. (2000) «Свидетельства изменений давления внешних факторов при МС1R» American Journal of Human Genetics 66: 1351—61. Связан с местным уровнем интенсивности ультрафиолетового излучения: Jablonski, N. and Chaplin, G. 261 Эпикантус — вертикальная кожная складка, прикрывающая медиальный угол глазной щели. — Прим. перев. 262 Основательное исследование изменений формы черепа у людей современного типа см. Lahr, M. (1996) «Эволюция многообразия типов современного человека: исследование изменений черепа» (Cambridge University Press); о характерных чертах внешности монголоидов и обзор монголоидов северного и южного типов см. там же, cc. 316—20. 263 В физическом и антропологическом отношении гораздо ближе к первым племенам-выходцам из Африки: Bulbeck, D. (1999) «Современные биологические и антропологические исследования негритосов Юго-Восточной Азии» SPAFA Journal 9(2): 15—22. Близкие к древнейшим черепам африканского и ближневосточного типа, возраст которых составляет 100 тысяч лет: Brown, P. (1999) «Первые люди современного типа в Восточной Азии? Новый взгляд на черепа Верхняя пещера 101, люйцзяньский и Минатогава 1» в K. Omoto (ed) «Междудисциплинарный взгляд на происхождение японцев» (International Research Center for Japanese Studies, Kyoto), cc. 105—30; см. также Bulbeck, указ. изд. 264 Bulbeck, указ. изд. 265 Сравнительную дискуссию см. в Lahr, указ. изд. 266 Ulijaszek, S. (2001) R.Martorell and F.Haschke (eds) «Питание и рост» Nestle Nutrition Workshop Series, Pediatric Program, Vol. 47 (Lippincott Williams & Wilkins, Philadelphia) cc. 1 — 15, Ulijaszek, S. (1994) «Вариации численного состава представителей предподростковой возрастной группы у разных народов» European Journal of Clinical Nutrition 48 (suppl. 1): S5—S14. 267 Уменьшение величины черепа у австралоидов: Lahr, указ. изд. Древнейшие ископаемые останки черепов, найденные в Австралии, принадлежали к «стройному», а не к «крепкому» типу. Другая причина, позволяющая считать черепа австралоидов несколько «изменившимися»: о других причинах, по которым современные аборигены Австралии по внешнему облику отличаются от первых переселенцев, см. в примечании 6 к главе 4. 268 Любопытно, что излюбленный мотив знаменитых японских нэцке (мини-пластики) — фигурки Фукурокудзи, божества-покровителя ученых, которого было принято изображать с гипертрофированно удлиненным черепом. Может быть, в этом отражена память о характерных длинных черепах исчезнувших древних этносов? — Прим. перев. 269 Племя тьерра дель фуэган: Lahr, указ. изд. Предками айнов: там же. 270 Имеется в виду Нельсон Мандела, первый президент Южно-Африканской Республики, избранный всенародным голосованием после падения диктатуры белого меньшинства. — Прим. перев. 271 Заметного сокращения расстояния между лбом и затылком: там же. Наиболее четко прослеживается на черепах людей эпохи неолита: Ishida, H. and Dodo, Y. (1996) «Морфология строения черепа у жителей Сибири и Восточной Азии» в A.Takeru and E.Szathmary (eds) «Расселение монголоидов в доисторическую эпоху» (Oxford University Press) cc. 113—24; Hanihara, Т. (2000) «Плоская форма лица у представителей крупнейших этнических групп» American Journal of Physical Anthropology 111: 105—34. 272 Конкретная дата появления в Китае первых людей современного типа: см. дискуссию о датировке люцзяньского черепа временем 68 тысяч лет тому назад в главе 4; кроме того, дискуссию о датировке люцзяньского черепа и черепа 101 из Верхней пещеры см. в Brown, указ. изд. Технологии эпохи Верхнего палеолита и останки человека современного типа, найденные на стоянке Салавасу в Северном Китае, относятся к периоду между 35 и 50 тысяч лет тому назад; см. Chen, С. and Olsen, J.W. (1990) «Китай в эпоху Последнего ледникового максимума» в C.Gamble and O.Soffer (eds) «Мир 18 000 лет тому назад» Vol. 1 (Unwin Hyman, London) сс. 276—95. Возраст самых древних в мире останков бесспорно монголоидного типа: в Brown, указ. изд. Эта изолированная находка вызывает целый ряд вопросов: описаны под названием «часть лобной кости, идентифицируемая как объект, имеющий сходство с восточноазиатским типом, что объясняется плоской верхней носовой областью» в Wolpoff, М. (1999) «Палеоантропология» (McGraw-Hill, Boston), с. 740; а также рассматриваются В. Алексеевым: Alekseev, V. (1998) «Физические особенности гоминидов эпохи палеолита в Сибири» в кн. AP.Derevjanko (ed) «Человек эпохи палеолита в Сибири: новые открытия и трактовки» (University of Illinois Press, Urbana). Необходимо отметить, что некоторые другие черепа древних обитателей Восточной Азии имеют, так сказать, статус «премонголоидов». Так, минатаговский череп с о. Окинава (возраст — 16 600—18 500 лет) имеет характерно плоскую переносицу. Эти черепа во многих отношениях (см. дискуссию в Brown, указ. изд.) отличаются от черепов монголоидов современного типа. Таким образом, хотя плоская переносица может считаться важным отличительным признаком, не вполне ясно, насколько она характерна для монголоидов. 273 Сохранение некоторых архаических особенностей скелета у современных европеоидов: Lahr, указ. изд. Европейцев тоже можно поставить в один ряд с обитателями прибрежной полосы Индийского океана (включая жителей Южной Индии, негритосов племени семанг (Малайя), и коренное население Новой Гвинеи), сохранившими куда больше архаических африканских особенностей строения зубов: Rayner, D. and Bulbeck, D. (2001) «Морфология строения зубов аборигенов племени орангасли на Малайском полуострове» в кн. М. Hennenberg (ed) «Причины и следствия вариаций типов человека» (Australasian Society for Human Biology, The University of Adelaide) cc. 19—41. И черепа: в одном из недавних краниометрических исследований было выявлено наличие признаков, характерных для черепов жителей Андаманских островов и народа семанг, тамилов Южной Индии и аборигенов побережья Новой Гвинеи, что свидетельствует о ряде наследственных признаков строения черепов и зубов — Bulbeck, указ. изд. 274 Коренного населения Малайского полуострова: известного как протомалайцы; этот постулат основан на старой теории о проникновении в этот регион двух волн миграции, см. Carey, I. (1976) «Оранг асли: аборигенные племена полуострова Малайзия» (Oxford University Press, Kuala Lumpur). Различные архаические этнические группы Тихоокеанского побережья, такие, как аборигены айны в Японии: Hanihara, Т. et al. (1998) «Место айнов о. Хоккайдо (Северная Япония) среди народов мира» International Journal of Circumpolar Health 57: 257—75, с. 273—4. 275 O происхождении сундадонтности см. Hanihara (1992), указ. изд. Толстые кости аборигенов Австралии и изменения строения зубов и черепов у них потенциально можно объяснить следствием второй волны колонизации Австралии выходцами из Юго-Восточной Азии в последние 20 тысяч лет (Wolpoff (1999), указ. изд., с. 740). Бердселл указывает на возможность трех ветвей колонизации, проникших в Австралию: первая из них — «океанские негритосы», или барринейцы, представителями которых были вымершие тасманцы, вторая — муррайанцы, близкие к айнам, а третья — карпентарийцы, близкие к веллоидам — Birdsell, J.B. (1977) «Пересмотр парадигмы для первых жителей Великой Австралии» в кн. J.Allen et al. (eds) «Сунда и Сахул» (Academic Press, London), cc. 113—67. Существует целый ряд археологических и генетических свидетельств в поддержку гипотезы о гораздо более позднем проникновении во время голоценового периода, однако культурологическое влияние было более мощным, чем генетическое, которое можно считать малозначительным — С. Оппенгеймера — Oppenheimer, S.J. «Эдем на Востоке» (1998, Weidenfeld & Nicolson, London), cc. 203—4. Хотя гипотеза о второй волне колонизации Австралии сегодня не в моде, и Бердселл и Уолпофф находятся в явном меньшинстве, я убежден, что данную гипотезу не следует сбрасывать со счетов и что генетические данные могут предложить аргументы в поддержку гипотезы о том, что заселение происходило по нескольким маршрутам. 276 Сундадонтность и синодонтность — достаточно искусственные критерии классификации некоего континуума. См. с одной стороны дискуссию в кн. Hanihara, Т. (1992) «Строение зубов у негритосов, аборигенов Австралии и «протосундадонтов»: коренное население Восточной Азии, V» American Journal of Physical Anthropology 88: 183—96; см. также Scott, G.R. and Turner II, C.G. (1997) «Антропология строения зубов у современного человека» (Cambridge University Press). Rayner and Bulbeck, (указ. изд.), с другой стороны, со всей определенностью указывают, что деление на сундадонтность и синодонтность является искусственным. Они указывают на клинальный континуум, считая, что сундадонтность занимает промежуточное положение между недиффренцированным околоиндийскоокеанийским типом и синодонтностью. В этой точке зрения есть место для клина Южной и Северной Азии. Исследователи также утверждают, что между этими народами Тихоокеанского побережья существуют значительные различия. Но, хотя я согласен с Рейнером и Балбеком в этом вопросе, по соображениям простоты я продолжаю пользоваться терминами «сундадонтность» и «синодонтность». 277 Адаптационные теории — теории, делающие главный акцент на приспособлении биоорганизмов или экосистем к условиям внешней среды — Прим. перев. 278 Географическое деление монголоидов на северян и южан становится еще более заметным при рассмотрении физических (и генетических) факторов: Yong-Gang Yao et al. (2002) «Филогеографическая дифференциация митохондриевой ДНК у китайцев-ханьцев» American Journal of Human Genetics 70: 635—51. Изучение динамики роста у детей в Китае показало, например, что дети жителей Гонконга (Южный Китай), несмотря на то что питались они гораздо лучше детей из континентального Китая: Leung, S. (1990) «Состояние здоровья детей-китайцев на о. Гонконг, обусловленное факторами оксологии и питания: от рождения до двух лет» MD Thesis, Chinese University of Hong Kong. Можно подумать, что малорослость и хрупкость — это отличительные черты монголоидов. Если северные монголоиды представляют собой ветвь южных, мы вправе были бы ожидать, что они скорее малорослые и хрупкие, нежели крупные и крепкокостные по сложению, но необходимо иметь в виду, что представители северных монголоидов смешивались с этническими группами Центральной Азии, связанными с жителями Западной Евразии, как об этом будет сказано ниже в данной главе. 279 Другая теория [адаптации к холоду]: Ishida and Dodo, указ. изд.; см. также Con, C.S. «Происхождение рас» (Alfred A.Knopf, New York); Akazawa, Т. (1996) «Введение: эволюция человека, расселение и принципы адаптации» в T.Akazawa and E.Szathmary (eds) «Расселение монголоидов в доисторическую эпоху» (Oxford University Press); и Guthrie, D. (1996) «Мамонтовая степь и происхождение и расселение монголоидов» в «в Т.Акаzawa and E.Szathmary (eds) «Расселение монголоидов в доисторическую эпоху» (Oxford University Press). Гатри увлеченно описывал: см. Guthrie, указ. изд. 280 Hanihara (2000), указ. изд., с. 117. 281 Ishida and Dodo, указ. изд. Альтернативная точка зрения [см. илл. 8 в Hanihara (2000)], заключается в том, что эти группы сохранили наследственные особенности строения Homo sapiens, в частности, плоское лицо, тогда как верхняя часть черепа, особенно лоб, резко сузилась, а величина челюстей также резко уменьшилась. 282 См. Montagu, А. «Рост в молодости» (McGraw-Hill, New York) с. 40; см. также Gould, S.J. (1977) «Онтегения и филогения» (Belknap Press, Cambridge, MA). Те, кто интересуется недавно изданными серьезными научными работами по этой тематике о взаимодействии — как позитивном, так и негативном — неотении и эволюции человека, см. Bemporad, J.R. (1991) «Слабоумие как крах неотении» Theoretical Medicine 12: 45—51; Crow, T.J. (2002) «Половой отбор, синхронистичность и гомологические гены X-Y: Могла ли Y-хромосома быть видообразующей для Homo sapiens?» Proceedings of the British Academy 106: 197—216. Несколько эвристических обзоров можно найти на сайтах www.neoteny.org/a/stephenjgould.html и www.humanevolution.net/a/asienoriental.html. 283 Шелковый путь появился на карте Азии за добрую тысячу лет до Марко Поло. Известно, что еще в I в. до н.э. по нему уже доставлялись шелка из Китая в Европу и в первую очередь — в Рим. Шелковый путь делился на несколько ветвей. В IX—XI вв южную ветвь, которая вела в Византию, контролировал Арабский халифат, а северную, которая вела в страны Западной Европы через северное Причерноморье, держал под контролем Хазарский каганат. — Прим. перев. 284 Сегодня — Синьцзян-Уйгурский автономный район КНР. — Прим. перев. 285 Chou, А. «Миграция ранних гоминидов в эпоху плейстоцена» paper presented at the Paleoanthropology Society Annual Meeting, 27—28 March 2001, Kansas City, Missouri. 286 В начале 1950-х гг государство Тибет было оккупировано войсками Китая и насильственно включено в состав КНР. — Прим. перев. 287 Появилась в этих местах около 43 тысяч лет назад...: Goebel, T. et al. (1993) «Датировка времени перехода от технологии Среднего палеолита к технологии Верхнего палеолита на стоянке Кара-Бом» Current Anthropology 34: 452—8. Переход к Верхнему палеолиту на всей территории Южной Сибири: Goebel, Т. and Aksenov, M. «Датировка по радиоуглеродному методу с ускорителем памятников начала Верхнего палеолита в Юго-Восточной Сибири» Antiquity 69: 349—57. 288 Otte, M. and Derevianko, A. (2001) «Ауриньякская культура на Алтае» Antiquity 75: 44—8; Kozlowski, J.K. and Otte, M. (2000) «Формирование ауриньякской культуры в Европе» Journal of Archaeological Research 56: 513—33; Otte, M. et al. (2000) «История населения Европы по данным археологии» в кн. С. Renfrew and К. Boyle «Археогенетика: ДНК и история заселения Европы в древнейшие времена» (McDonald Institute for Archaeological Research, Cambridge), cc. 139—41; Otte, M. (2003) «Памятники ауриньякской культуры в Азии» (готовится к печати); автор, между прочим, цитирует статью Olszewski, D.I. and Dibble, H.L. (1994) «Памятники ауриньякской культуры в горах Загрос» Current Anthropology 35(1): 68—75. 289 Goebel et al., указ. изд. Otte, M. and Derevianko, А., указ. изд. 290 Sine qua non (лат.) — обязательное условие. — Прим. перев. 291 Находки в пещерах Алтая (Россия), в частности — в пещере Кара-Бом, свидетельствуют о наличии традиционных техник Среднего палеолита...: Otte, М. and Derevianko, A. (1996) «Техники обработки камня у людей эпохи палеолита на Алтае (Сибирь)» Anthropologie et Prehistoire 107: 131—43. Постоянное накапливание технических навыков, позволившее совершить прорыв в технологии изготовления орудий: Otte, M. and Derevianko, A. (1996). Свидетельства присутствия человека современного типа на Алтае: Goebel, Т. and Aksenov, указ. изд., с. 356. См. также М. Goebel et al., указ. изд. Otte, M. and Derevianko, A. (2000) «Археологические исследования, осуществленные совместной российско-монгольско-американской экспедицией в Монголии в 1997—1998 гг.» (Институт археологии и этнографии, Новосибирск), сс. 161, 168. 292 Возраст традиционных технических навыков Среднего палеолита: Blackwell, B.A.B. et al. (1998) «Датировка по методу электронного резонанса (ESR) палеолитической стоянки в Цаган-Агуй, Монголия» Proceedings of the 31st International Symposium on Archaeometry, 27 April - 1 May 1998, Budapest. Существовали две раздельные микролитические традиции эпохи Верхнего палеолита: Tang Hui Sheng (1995) «Очаги изготовления каменных орудий на Цзиньхай-Тибетском плато» The Artefact 18: 3—11. 293 Soffer, О. (1993) «Адаптации эпохи Верхнего палеолита в Центральной и Восточной Европе и взаимодействие человека с мамонтом» в O.Soffer and N.Praslov «От Костенки до Кловиса: адаптации палеоиндийских ветвей в эпоху Верхнего палеолита» (Plenum, New York), cc. 33—49. 294 Cavali-Sforza, L et al. (1994) «История и география генов человека» (Princetown University Press, Princetown) cc. 223-38. 295 Строго говоря, основное территориальное ядро исторической Китайской империи («древние» полумифические владыки, правившие, по преданию, по 2000—2500 лет, естественно, не в счет) сложилось в правление ее основателя, Цинь Шихуанди (III в до н.э.), а этноним «хань» стал употребляться для обозначения китайцев начиная с правления династии Хань (II в до н.э. — I в н.э.). Сегодня на долю ханьцев приходится более 92% населения КНР. — Прим. перев. 296 Chang Wing-hoi (1995) «Посвятительные имена в генеалогиях хакка: религиозная практика и ее упадок» в D.Faure and H.Siu (eds) «Вдоль по земле: связи между территориями в Южном Китае» (Stanford University Press) cc. 65—82. Ряд ученых, по не вполне понятным причинам, до сих пор поддерживают миф об этнической однородности населения на всей территории Китая (см., например, Yuan-Chun Ding et al. (2000) «Структура населения и история Юго-Восточной Азии» Proceedings of the National Academy of Sciences USA 97: 14003—6; переработанный вариант при участии тех же авторов, см. Yong-Gang Yao et al. (2002) «Филогеографическая дифференциация митохондриевой ДНК у китайцев-ханьцев» American Journal of Human Genetics 70: 635—51. 297 Su, B. et al. (1999) «Y-хромосомные свидетельства миграции людей современного типа на север, в Восточную Азию, во время последнего Ледникового периода» American Journal of Human Genetics 65: 1718—24. 298 Палеоклиматология — научная дисциплина, изучающая состояние климата и прочих метеорологических факторов в давние эпохи истории Земли — Прим. перев. 299 Палеоклиматологические исследования: см. карты на сайте http://www.esd.ornl.gov/projects/qen/NEW...MAPS/eurasia1.gif и /www.esd.ornl.gov/projects/qen/eurasia2.gif. Наличие генетического многообразия и поливариантности на юго-западе Центральной Азии: см. илл. 2 в Metspalu, E. et al. (1999) «Закавказье и распространение специфически кавказоидного типа митохондриевой ДНК» в кн. S.S.Papiha et al. «Многообразие геномов и его применение в генетике человека» (Kluwer Academic/Plenum Publishers, New York) cc. 121 — 134. Специальные карты на вебсайте Джонатана Адамса ESD ORNL /www.esd.ornl.gov/projects/qen/euras18k.gif и /www.esd.ornl.gov/ projects/qen/euras(2.gif. См. также археологические свидетельства, представленные в следующей главе. 300 Группы А и X представляют собой первое поколение потомков N/Насрин, тогда как В и F происходят от Насрин через Рохани. Группы С, D, E, G и Z являются прямыми потомками М/Манью (см. илл. 5.9 и 5.10). Первое полное описание групп A-G, показанных на одном древе, было представлено в Torroni, A. et al. (1994) «Анализ митохондриевой ДНК на Тибете: его использование для установления происхождения тибетцев и их способности адаптироваться к условиям высокогорья» American Journal of Physical Anthropology 93: 189—99. О первоначальном наименовании группы X см. Forster, P. et al. (1996) «Происхождение и эволюция вариаций мтДНК у коренных народов Америки: пересмотр традиционной оценки» American Journal of Human Genetics 59: 935—45. Другая группа, Y, встречается почти исключительно на севере, у народности нивхов на о. Сахалин; эта группа очень молода и поэтому находится на периферии дискуссии (см. ниже). О группах Y и Z, а также о переоценке структуры древа азиатской мтДНК см. Kivisild, Т. et al. (2002) «Новые ветви и отрасли на древе мтДНК Восточной Азии» Molecular Biology and Evolution 19(10): 1737— 51; Yong-Gang Yao et al. (2003), указ. изд.; и илл. 2 в Forster, P. et al. (2003) «Эволюция мтДНК у аборигенов Азии и Папуа» в P.Bellwood and C.Renfrew (eds) «Исследования гипотезы о распространении сельского хозяйства и языка» (McDonald Institute for Archaeological Research, Cambridge), cc. 89—98. Главная задача заключалась в том, чтобы выявить различия между северной и южной генетическими линиями у монголоидного населения. С и Z — типы, связанные с группой М (о недавно выявленной «родительской» группе М8 см. Yong-Gang Yao et al. (2003), указ. изд.), и встречаются они почти исключительно в Северной Азии (а С также в Северной и Южной Америке), а не у южных монголоидов. Тип А, хотя он и распространен на северо-востоке Китая, в северо-восточной Сибири и Северной Америке (Kolman, C.J. et al. (1996) «Анализ митохондриевой ДНК у монголоидного населения и его использование для установления корней происхождения групп-родоначальников Нового Света» Genetics 142: 1321—34), отличается весьма частой встречаемостью и многообразием в южном и юго-западном Китае, на Тибете и Синьцзяне (см. Yong-Gang Yao et al. (2003), указ. изд.), но совершенно отсутствует в Индии и Западной Евразии (см. Kivisild, Т. et al. (1999) «Место индийских вариантов митохондриевой ДНК в глобальной системе материнских генетических линий и заселении Старого Света» в кн. S.S.Papiha et al. «Многообразие геномов и его применение в генетике человека» (Kluwer Academic/Plenum, New York) cc. 135—52. Таким образом, этнический источник в юго-западном Китае, с последующим продвижением по Янцзы на Тибет, представляется гораздо более вероятной прародиной А, чем прочие источники в Индии или Северной Азии. Группа Y, дочь N, выявлена только в Северо-Восточной Азии (Stariovskaya, Y.B. et al. (1998) «Многообразие мтДНК у чукчей Сибири: возможности применения для изучения генетической истории Древней Берингии и заселения Нового Света» American Journal of Human Genetics 63: 1473—91); Torroni, A. et al. (1993) «Вариации мтДНК у коренных жителей Сибири свидетельствуют о несомненной генетической близости с туземным населением Америки» American Journal of Human Genetics 53: 591—608). Однако возраст группы Y — всего лишь около 5000 лет, и она происходит от южноазиатской ветви N — N9. Распространение X, еще одной «дочери» N, ограничено европейцами и американцами (Brown, M. et al. (1998) «Гаплогруппа X мтДНК: древнее связующее звено между Европой/Западной Азией и Северной Америкой?» American Journal of Human Genetics 63; 1852— 61; кроме того, есть единичные сообщения о выявлении X у аборигенов Сибири (см. ниже). Хотя любая из этих трех линий (С, X и Z) теоретически могла стать характерной чертой генетических линий монголоидов, все эти линии полностью отсутствуют у южных монголоидов, но зато встречаются у других этнических групп, что делает их присутствие еще менее вероятным. В последнее время к этим группам присоединилось еще несколько кладов, выявленных в Китае: три из них происходят от М (М7, М9 и М10), а четвертый — от N (N9, примерный возраст 64300 ... 20 тысячи лет; является наследственным для Y): см. Yong-Gang Yao et al., указ. изд. Две азиатские «внучки» N (В и F) — единственные известные науке восточноазиатские потомки линии R/Poхани (не считая вновь описанного древнего клада, R9, возраст которого — 81 000 ... 24 600 лет; см. Yong-Gang Yao et al., указ. изд., и который является наследственным для F на юге и поэтому получил название F4 в Hill, С. et al. «Вариации митохондриевой ДНК у племени оранг асли на Малайском полуострове» American Journal of Human Genetics (готовится к печати); см. илл. 5.5). В и F отличаются наибольшим разнообразием и наивысшей встречаемостью у южных монголоидов Юго-Восточной Азии (Torroni, A. et al., указ. изд., Yong-Gang Yao et al., указ. изд.). Они выявлены также к северу от Гималаев в Центральной Азии (илл. 2 в Metspalu et al., указ. изд.), в Монголии (Kolman et al., указ. изд.), а также на Тибете, в Корее и Японии (Torroni, A. et al. (1994), указ. изд.; Horai, S. et al. (1996) «Полиморфизм мтДНК у населения Восточной Азии» American Journal of Human Genetics 59: 579—90). В то же время эти линии не встречаются на Крайнем Севере Сибири и в субарктическом регионе. Из этих южных линий В также выявлена в Америке, хотя опять-таки не в субарктическом регионе (см. главу 7). Так как линии В и F присутствуют по обе стороны Гималайского хребта, они являются потенциальными претендентами на роль линий-родоначальниц монголоидов, однако вопрос о том, присущи ли они всем монголоидам (линия F в Америке отсутствует), остается открытым. Мы рассмотрели три северноазиатские линии и три линии из Южной Азии, так что у нас остаются линии D, Е и G. Эти «дочери» М/Манью сохраняют потенциальную связь со своими истоками (дело в том, что они все восходят к М и имеют общую мутацию в быстрой (нестабильной) точке 16362 — см. Kivisild et al. (1999, 2002), указ. изд.). Линии D и G широко распространены на всей территории расселения монголоидов, а также в Индокитае. Уникальные локальные варианты D в большом количестве встречаются в Китае, Японии, Монголии, на Тибете, в Корее, Центральной и Северо-Восточной Азии, а также в Северной и Южной Америке (Torroni et al., (1994), указ. изд.; Torroni, A. et al. (1993) «Черты сходства с азиатами и континентальное распространение четырех ветвей-родоначальниц мтДНК коренных жителей Америки» American Journal of Human Genetics 53: 563—90; см. Kolman et al., указ. изд.; Kivisild et al. (1999, 2002), указ. изд. Хотя линия D и встречается в Южном Китае, она не характерна для Юго-Восточной Азии (Torroni et al. (1994), указ. изд.). Уникальные варианты линии G выявлены в Центральной Азии (см. илл. 2 в Metspalu et al., указ. изд.), на Тибете (Torroni et al., (1994), указ. изд.), у айнов (Horai et al., указ. изд.) и в Северо-Восточной Азии (Torroni et al., (1994), указ. изд.). Группа Е имеет свою собственную уникальную подгруппу на островах Юго-Восточной Азии, а также встречается на Тибете (типы Торрони 83, 89, 94, 104, 106, 109 и 119; последний идентифицирован неточно в связи с присутствием RFLP в точке 16517, Torroni et al. (1994), указ. изд.; илл. 2 в Metspalu et al., указ. изд.). Вновь описанный клад, М7 (возраст — 61 ... 20 тысяч лет), как и D, широко распространен в прибрежных районах Восточной Азии, но, подобно Е, вектор его распространения направлен в Юго-Восточную Азию: см. Kivisild et al. (2002), указ. изд.; Yong-Gang Yao, указ. изд. 301 Данные для Центральной Азии — илл. 2 в Metspalu et al., указ. изд., для Тибета — Torroni et al., (1994), указ. изд., для Монголии — Kolman et al., указ. изд.; для Китая и Кореи — Torroni et al., (1994), указ. изд. 302 Возраст четырех из этих ветвей в Монголии: Kivisild et al. (1999), указ. изд. (Обратите внимание, что существует возможность переоценки многообразия линий, влияющей на локальные показатели возраста этих линий в Монголии.) Средний возраст тех же линий на юге, в Южном Китае: табл. 3 в Yong-Gang Yao, указ. изд. Обратите внимание, что в Yong-Gang Yao, указ. изд., показано, что возраст индивидуальных ветвей D и G составляет 51—60 тысяч лет и что эти гаплогруппы совмещаются с М9/Е даже в более раннюю эпоху (Kivisild et al. (1999), указ. изд.). 303 Группа В, возраст которой в Азии составляет около 75 тысяч лет (74 600 ... 18 700 лет, Yong-Gang Yao, указ. изд.), обладает наивысшей встречаемостью у коренных жителей Юго-Восточной Азии, Океании (за исключением горцев Новой Гвинеи и аборигенов Австралии) и западного побережья Тихого океана. 304 О многообразии и древности В и F на юге см. Ballinger, S.W. et al. (1992) «Анализ митохондриевой ДНК жителей Юго-Восточной Азии свидетельствует о постоянном характере миграций древних монголоидов» Genetics 130: 139—52. Данные взяты также из Fucharoen, G. et al. (2001) «Аспекты полиморфизма митохондриевой ДНК в Таиланде» Journal of Human Genetics 46: 115—25; и Oota, H. et al. «Крайняя однородность мтДНК у коренного населения континентальной Азии» American Journal of Physical Anthropology 118: 146—53; см. также Kivisild et al. (1999), указ. изд., Yong-Gang Yao, указ. изд. Группы В (примерный возраст — 74 600 ... 18 700 лет) и R9(F4) (примерный возраст — 81 000 ... 24 600 лет), которая является наследственной для F на юге (см. выше, примечание 33). Является вновь идентифицированная гаплогруппа «npe-F»: см. Hill, С. et al., указ. изд. См. илл. 5.5. Вновь описанная гаплогруппа М7 отражает данные по В и F. Возраст М7 — 61 тысяча лет, а ее наиболее ранняя ветвь, М7b, является характерной чертой жителей Вьетнама — см. Kivisild et al. (2002), указ. изд. 305 Возраст двух основных подгрупп В: подгруппа В1, являющаяся древнейшей в Монголии (40 500 лет), представляет собой тип, доминирующий в Юго-Восточной Азии, в Тихоокеанском бассейне и в Северной и Южной Америке. Тип В1 выявлен также на Тибете — см. Kivisild et al. (2002), указ. изд; Torroni et al., (1994), указ. изд. Дифференциация типов В1 и В2 производится соответственно по точкам 16217 (согласно официальной классификации — В4) и 16243/16140 (сегодня, согласно классификации Kivisild et al. (2002), указ. изд., — В5b) в работе Kolman et al., указ. изд. Более молодой из этих типов, В2 (его возраст в Монголии — около 33 500 лет), в большей мере, чем В1, распространен на западном побережье Тихого океана (Восточный Китай, Корея, Япония) — см. Horai, S. and Hayasaka, К. «Различия между интраспецифическими нуклеотидами в обширной некодированной области митохондриевой ДНК человека» American Journal of Human Genetics 46: 828—42; Nishimaki, Y. et al. (1999) «Последовательный полиморфизм в зоне HV1 мтДНК у японцев и китайцев» Legal Medicine 1: 238—49; Horai et al., указ. изд.; Seo, Y.B. et al. (1998) «Последовательный полиморфизм в контрольной зоне митохондриевой ДНК у японцев» Forensic Science International 97: 155—64. Оба подтипа В могли появиться еще в эпоху палеолита: илл. 2 в Metspalu et al., указ. изд. 306 Надо упомянуть и о трех линиях — D, Е и G, — частично связанных с Манью: линия D, занимающая доминирующее положение среди этих линий и имеющая возраст около 44 500 лет (Kivisild et al. (1999), указ. изд.), является наиболее распространенной в Сибири, присутствует в Южном Китае и в Северной и Южной Америке, но не встречается в Юго-Восточной Азии, напр. в Сундаленде (Torroni et al., (1994), указ. изд.). Линия D, правда, значительно реже, представлена в Индии (Kivisild et al. (1999), указ. изд.). Линии D и Е также распространены на западе Центральной Азии (илл. 2 в Metspalu et al., указ. изд.; о линии Е см. Ballinger et al., указ. изд.). Известно, что А-вариант линии Е, присутствующий в Корее, широко распространен в Юго-Восточной Азии, что увеличивает вероятность того, что предок этой группы был выходцем с юга. Кроме того, существует прямая связь между G и уникальной группой М на о. Семанг, свидетельствующая о наличии возможного южного источника происхождения этой группы (неколированная точка 5108 в Hill et al., указ. изд.) Ветви-сестры С и Z наиболее распространены: Kolman et al., указ. изд.; Bamshad, M. et al. (1999) «Генетические свидетельства происхождения представителя разных каст в Индии» Genome Research 11: 994—1004; Yong-Gang Yao, указ. изд.; Kivisild et al. (1999), указ. изд. С и Z наиболее распространены в Сибири и Северо-Восточной Азии ... Монголии, Тибете и Центральной Азии... вплоть до Турции: Kivisild et al. (1999), указ. изд. (С — только в Индии), Bamshad, M. et al. (1999). Возраст группы С в Монголии: Kivisild et al. (1999), указ. изд. 307 Есть сообщение о находке следов линии X в Южной Сибири: Derenko, M.V. et al. (2001) «Присутствие гаплогруппы X митохондриевой ДНК у алтайцев Южной Сибири» American Journal of Human Genetics 69: 237—41. Как минимум 30 тысяч лет назад: Brown et al., указ. изд. 308 Линии D/E, С и F представляют собой согласованные термины для YAP+, RPSAY/M216 и М89 см. Консорциум по изучению Y-хромосомы (2000) «Система обозначений для генеалогического древа бинарных гаплогрупп Y-хромосомы человека» Genome Research 12: 339—48. 309 В Восточной Индонезии, Австралии и Новой Гвинее: гаплотип 48 — в Восточной Индонезии и Новой Гвинее, а гаплотип 49 — в Австралии. Продвигались вдоль побережья Индийского и Тихого океанов вплоть до берегов Японии и Кореи: гаплотип 50. Сын-азиат, ставший родоначальником многих народов: М217 или гаплотип 52, способствовавший развитию гаплотипов 51 и 53 — все гаплотипы см. в Underhill, P.A. et al. (2001) «Филогеография бинарных гаплотипов Y-хромосомы и истоки происхождения современного человека» Annals of Human Genetics 65: 43— 62. Проникли в Монголию и Центральную Азию: Karafet, T.M. et al. (1999) «Источник(и) по азиатским предкам гаплотипов Y-хромосомы у родоначальников населения Нового Света» American Journal of Human Genetics 64: 817—31. Оказалась в Америке: Underhill, Р.А. et al. (2000) «Вариации последовательности развития Y-хромосомы и история человечества» Nature Genetics 26: 358—61. 310 Крайней границей их продвижения на север стала Корея: Karafet et al., указ. изд. В степях Монголии и на Алтае (Россия): данные см. там же; см. также данные в Bing Su et al. (1999) «Y-хромосомные свидетельства миграции людей современного типа на север, в Восточную Азию, во время последнего Ледникового периода» American Journal of Human Genetics 65: 1718—24. 311 Из общего числа 12 127 обследованных жителей островов Азии и Океании: М89 (согласованный термин для ветви F/Сиф) см. в Ке, Y. et al. «Африканские корни происхождения людей современного типа в Восточной Азии: история 12 000 Y-хромосом» Science 292: 1151—2. В этой работе сказано также, что в пробах ДНК у всех 12 127 жителей Азии выявлено присутствие характерной мутации Адама выходцев из Африки, отмечаемой у 99,99% всех неафриканцев на нашей планете. Как утверждают некоторые генетики: Мнение о том, что Сиф появился в результате более позднего исхода из Африки, высказано в Underbill et al., указ. изд. Сифитские черты внешнего облика у представителей древнейших этнических групп собирателей на прибрежной полосе Индийского океана: см. (а) М89 (согласованный термин для группы F) 23% и М95 (согласованный термин для группы О) 65%, соответственно, у аборигенов племени оранг асли на Малайском полуострове — см. данные в Bing Su et al. (2000) «Корни происхождения полинезийцев с точки зрения миграции Y-хромосом» Proceedings of the National Academy of Sciences USA 97: 8225—8; (b) M9 (согласованные термины для группы К, включая подгруппы О, L и Р) — у 100% жителей Больших Андаманских островов, тогда как азиатская YAP+ (Каин, или согласованный термин для группы D) — у 100% жителей других островов Андаманского архипелага; данные см. Thangaraj, К. et al. (2002) «Генетические аспекты у жителей Андаманских островов — вымирающих этнических групп» Current Biology (published online 26 November); (с) М9, у 98% австралоидных племенных групп в Индии — кенчу и койа — данные см. Kivisild et al. (2002), указ. изд.; (d) Kayser, M. et al. (2001) «Независимые линии Y-хромосом человека в пробах из Меланезии и Австралии» American Journal of Human Genetics 68: 173—90. 312 Ha долю самого Сифа пришлась примерно четверть вариантов Y-хромосомы, а на долю его сынов и потомков — большая часть остальных вариантов: о Сифе см. илл. 1 в Hammer, M.F. et al. (2001) «Иерархическая картина глобального многообразия Y-хромосомы человека» Molecular Biology and Evolution 18(7): 1189—1203; о представителях линий «сыновей» Сифа, гаплотипов 19—24, см. илл. 1, там же. 313 Hammer et al. (2001), указ. изд., гаплотипы 20—23, см. также илл. 1, там же. См. также главы 3 и 4. Согласованный термин для группы F/Сиф относится и к G-R в Консорциум по изучению Y-хромосомы ( 2002), указ. изд. 314 На долю Кришны приходится около 40% типов Y-хромосом: Underbill et al. (2001), указ. изд.; Hammer et al. (2001), указ. изд. Он появился на свет в Индии вскоре после первоначального исхода из Африки: Согласно одной из оценок (по «филогенетическому методу»), возраст непосредственного предка Кришны, линии М89 (Сиф), составляет 88 тысяч лет; возраст же подветви Поло (M17) в Индии составляет 51 200 лет (при использовании того же метода датировки). Впоследствии M17 мигрировала в Центральную Азию и Европу — см. табл. 3 в Kivisild, T. (2003b) «Генетика языка и распространение сельского хозяйства в Индии» в кн. P.Bellwood and C.Renfrew (eds) «Исследования гипотезы о распространении сельского хозяйства и языка» (McDonald Institute for Archaeological Research, Cambridge), cc. 215—222. Некоторые из них поселились в Пакистане и Индии: гаплотипы 90 и 91 в гаплогруппе, обозначаемой М11, см. Underbill et al. (2000), указ. изд. Гаплотипы 90 и 91, определяемые по M147 и М70, см. там же. третьих можно встретить только в Меланезии: гаплотипы 94—97, определяемые по M4G/ M5T/M9G, см. Kayser et al., указ. изд., а также Capelli С. et al. (2001) «Преобладающее генетическое наследие коренных жителей Юго-Восточной Азии и Океании, говорящих на языках австралонезийской группы» American Journal of Human Genetics 68: 432—43. Наконец, представители еще одного типа (ТАТ) встречаются только в Центральной Азии и Северо-Восточной Европе: ТАТб гаплотип 92, см. Underhill et al. (2000), указ. изд. 315 Ч ж э н X э (XV в.) — знаменитый китайский флотоводец и кораблестроитель династии Мин, организатор дальних морских экспедиций, в том числе — к берегам Аравии и Африки. Любопытно, что по вероисповеданию Чжэн Хэ был хуэйхуэй — так в Китае называли этнических китайцев, принявших ислам. Кстати сказать, именно этим объясняется чуждая китайцам страсть Чжэн Хэ к дальним странствиям, поскольку он, будучи мусульманином, просто обязан был совершить хадж в Мекку, находящуюся в Саудовской Аравии. — Прим. перев. 316 Хо Ши Мин (наст. имя Нгуен Аи Хак) — один из организаторов освободительного движения в Индокитае, создатель Компартии Вьетнама и первый президент ДРВ. В последние годы жизни удалился от дел и жил отшельником в хижине на островке, воплощая идеал конфуцианского совершенного мужа и буддийского святого. — Прим. перев. 317 Одна из этих ветвей, М175 или согласованный тип О... Хо: ветвь M175 см. Underhill et al. (2000), указ. изд. О/Но соответствует гаплогруппе О — см. Консорциум по изучению Y-хромосомы (2002). Хо, в свою очередь, делится на три ветви...: Одна из них осталась в Южном Китае, Индокитае и Юго-Восточной Азии...: М95. Вторая ... проникла в Южный Китай, причем наиболее высокая концентрация представителей этой ветви отмечена на о. Тайвань: M119 Третья ... достигла Японии, Кореи и Северо-Восточной Азии: Bing Su et al. (1999, 2000), указ. изд.; Karafet et al., указ. изд.; Underhill et al. (2000), указ. изд. 318 Потомки другого азиатского «сына» Кришны — Поло (М45): ветвь М45 в Underhill et al. (2000), указ. изд. Представители ветви Поло проникли в Центральную Азию из Пакистана: Консорциум по изучению Y-хромосомы (2002), указ. изд. Кетов и селькупов: Karafet et al., указ. изд. 319 О. Soffer and N. Praslov «От Костенки до Кловиса: адаптации палеоиндийских ветвей в эпоху Верхнего палеолита» (Plenum, New York). |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Вверх |
||||
|
|
||||